海生爬行动物
Marine reptiles

地球上自6亿年前出现动物以来,曾发生6次重大的生物灭绝事件,其中最著名的要算6500万年前的恐龙灭绝。但是规模最大、对生物发展影响最深远的一次是发生在2.5亿年前的二叠纪末期。这是一次超级大灭绝,97%的海洋生物消失,生态系统也彻底更新,成为地球历史从古生代向中生代转折的里程碑。但是,产生这一过程的原因至今仍是一个科学谜团。有人认为原因是超大规模火山爆发、海平面的下降、水化甲烷大规模释放以及彗星等天体撞击等,但是一直没有充分的证据。
专家通过对澳大利亚帕斯盆地岩芯的地层和浙江煤山岩石剖面开展生物分子化石的研究,得出2.5亿年前后全球发生过海洋硫化氢污染的初步结论。由于硫化氢的“毒害”,造成当时海洋和陆地生物的大面积灭绝。
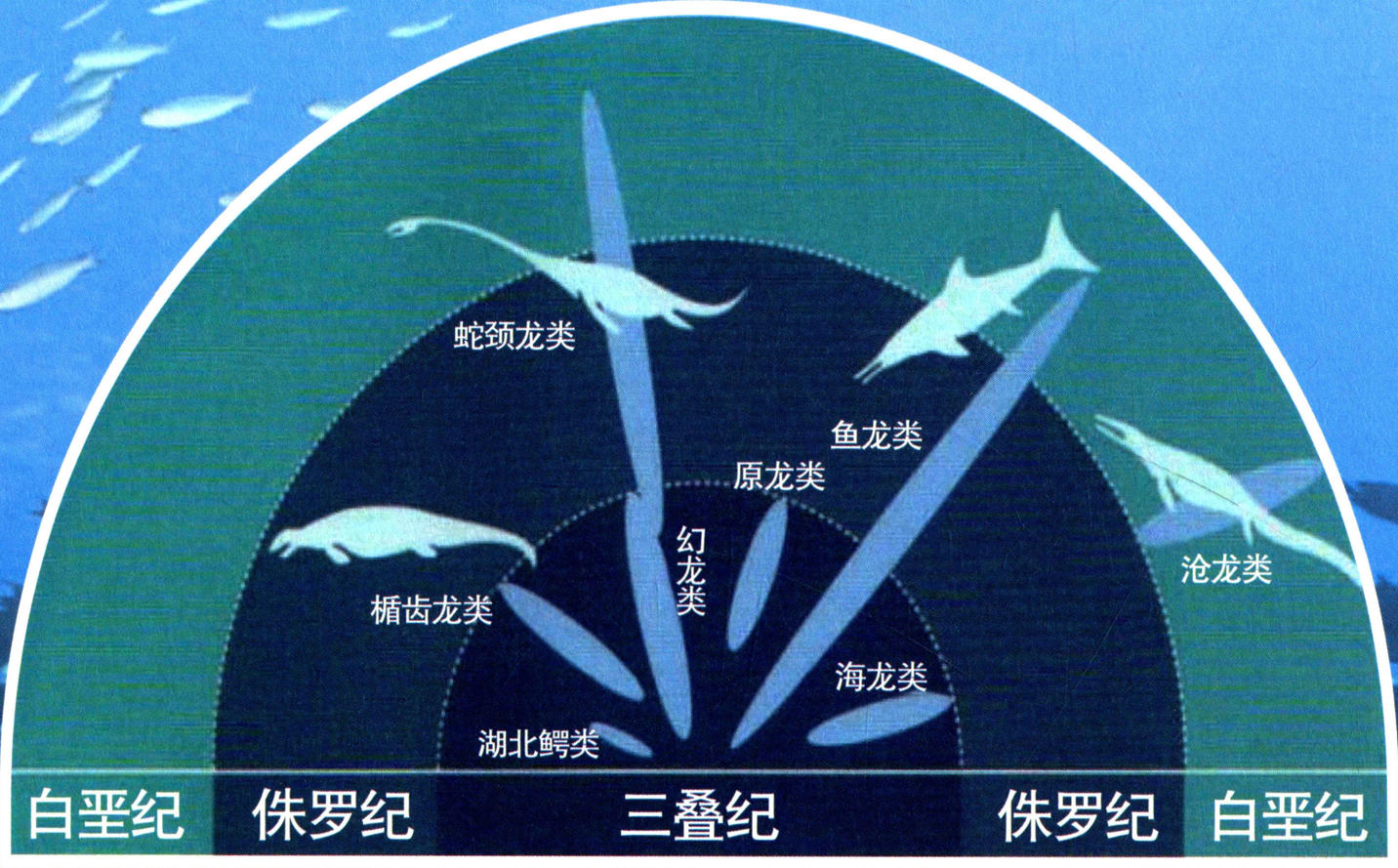
爬行动物分纪图
过了两千万年,地质历史进入三叠纪。由于全球性的大海侵,这时的贵州,一片汪洋大海。在深深浅浅的海域中,出现了形形色色的海生爬行动物。脊椎动物用了一亿多年时间,完成了从水生到陆生的转变;而从陆生又返回水中则要容易得多。海洋里的食物更为丰富多样,海洋里更适合爬行动物繁衍生息。于是,贵州这片海域成了全球海生爬行动物最发育地区之一。
海生爬行动物中,鳍龙类、海龙类和鱼龙类等先后在海洋里出现并逐步形成辐射发展。
在刚刚经历过一场大灭绝的生物界,海生爬行动物爆发式的出现,可能是三叠纪最出彩的篇章了!
而今的贵州,完好地保存着三叠纪浅海的石灰岩台地和深海沉积盆地的地质遗迹,埋藏着丰富的三叠纪古生物化石,被誉为“沉积岩王国”和“古生物王国”,记录着有待解密的二叠纪生物大灭绝和三叠纪生物大复苏的完整信息。
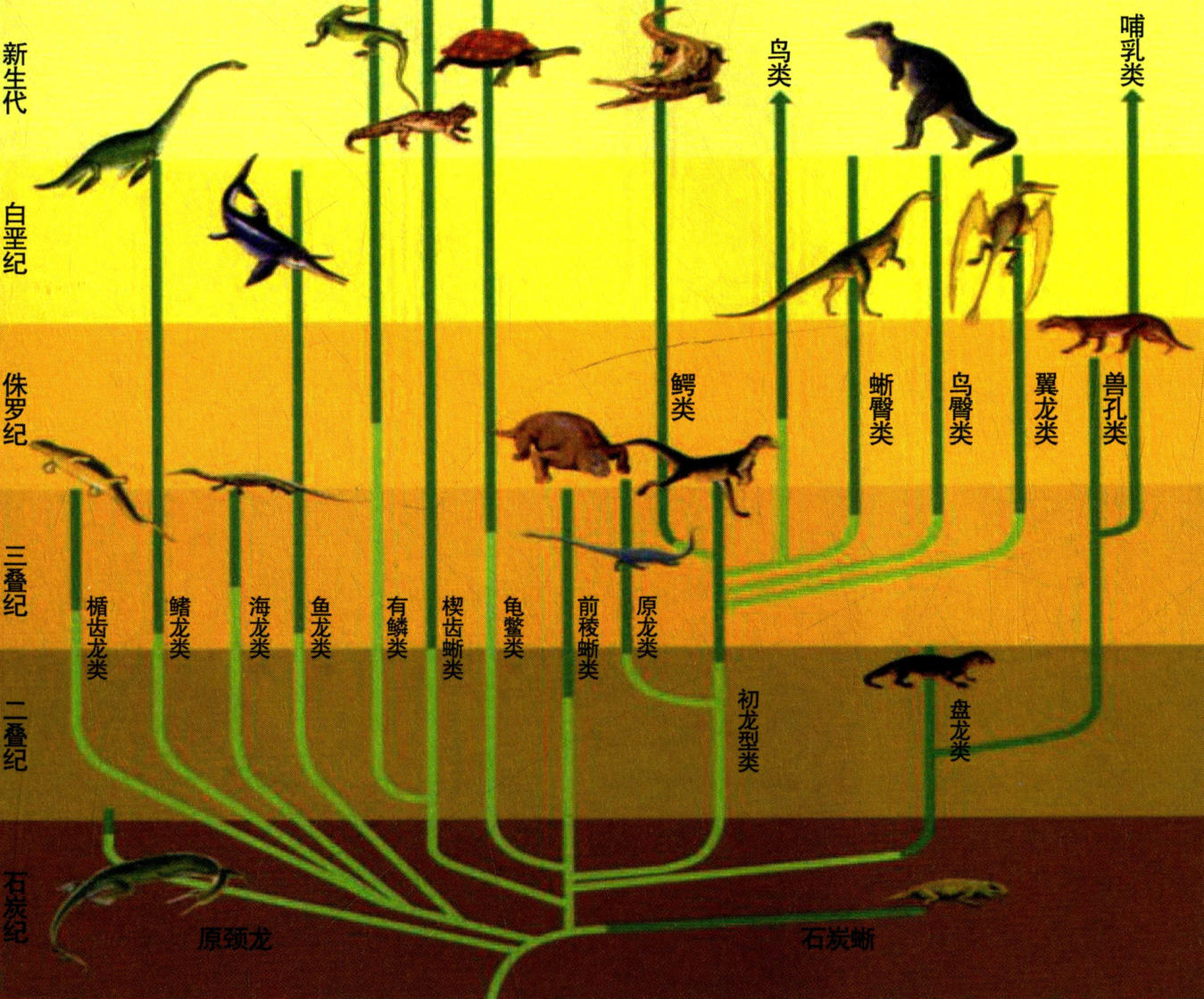
爬行动物演化图

始鳍龙类
Eosaropterygians
始鳍龙类是指楯齿龙以外的鳍龙类的成员,是中生代海洋中的捕猎者,包括三叠纪近岸生活的肿肋龙类、幻龙类、纯信龙类以及由幻龙类的一支发展而来的蛇颈龙类。

一、肿肋龙类
Pachypleurosaurids
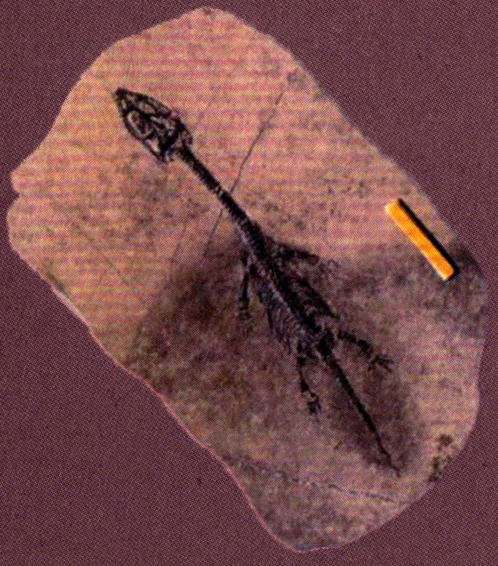
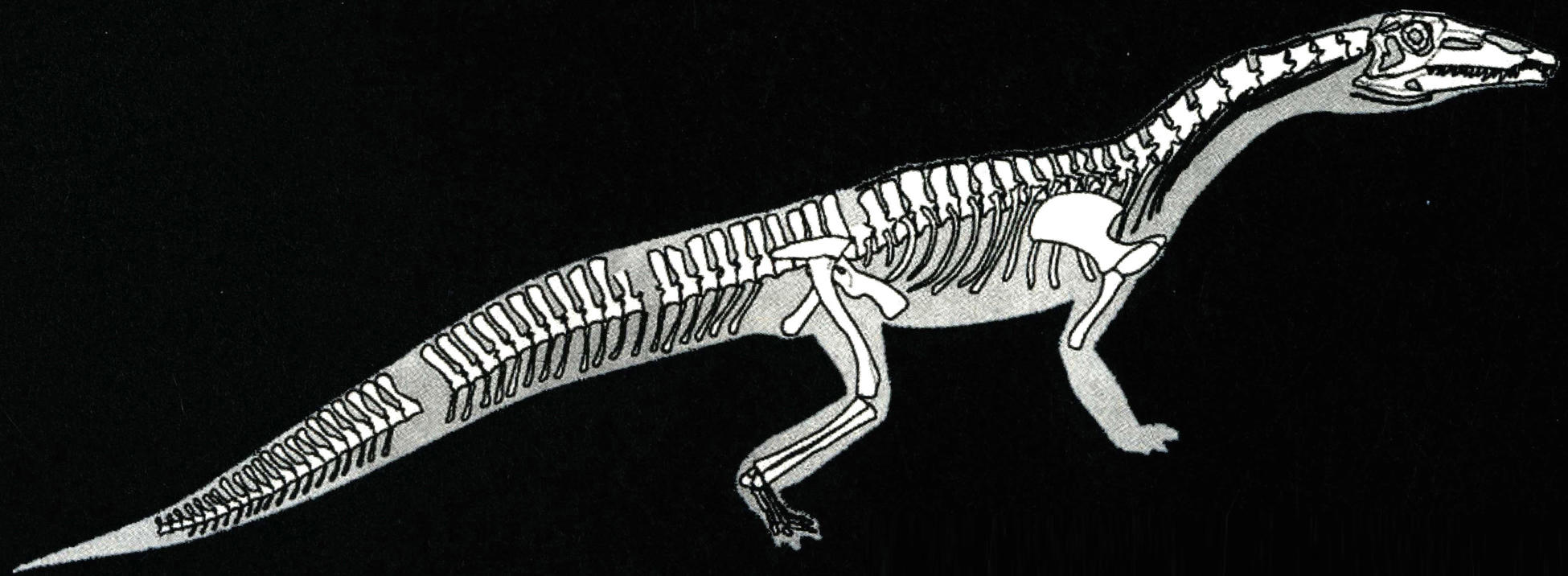
肿肋龙类是小型、轻巧,状似蜥蜴的爬行动物,体长大多不到1米,有的种类成体仅长20~30厘米。肿肋龙类的四肢只是初步适应了水生生活,所以只能在近岸的浅海区捕食小鱼和其他海洋生物。在贵州发现的肿肋龙类化石有胡氏贵州龙(Keichousaurushui)和细齿乌蒙龙(Wumengoasurus delicatomanlibularis)。
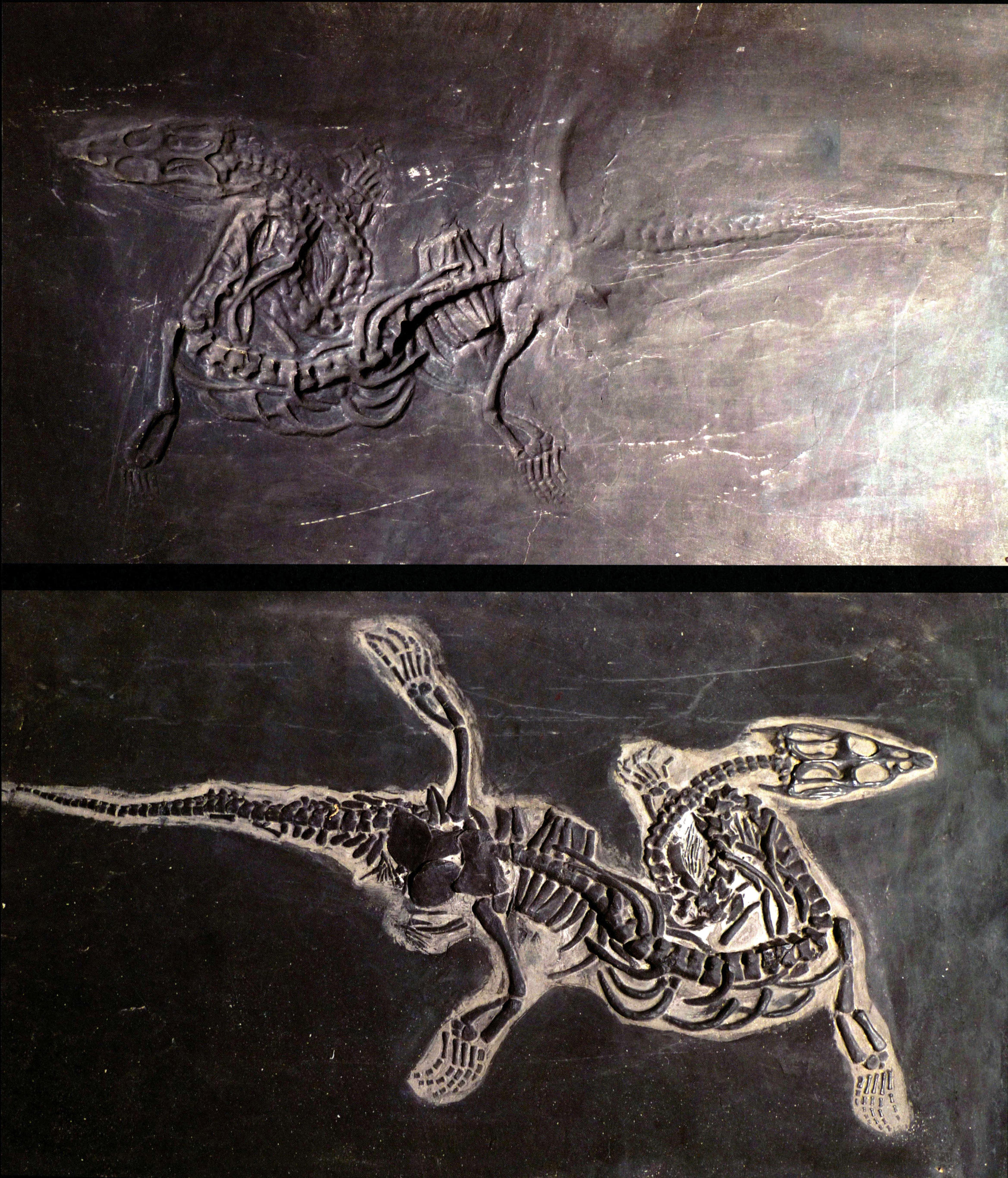
胡氏贵州龙的发现及其意义
1957年7月,中国地质部陈列馆(现中国地质博物馆)胡承志先生到贵州进行地质调查。在兴义市顶效镇大寨村浪幕山,胡先生发现了一些类似于蜥蜴的小型爬行动物化石,并采集了8块带到北京,交给中国科学院古脊椎馆动物与古生物研究所杨钟健教授;其后贵州省博物馆曹泽田等两次来到同一地点采集了另外7块标本带到北京。杨钟健对这些标本进行了详细研究,识别出这是一种从未定名的肿肋龙化石,是在中国乃至东亚地区的首次发现,并将其命名为胡氏贵州龙。

鳍龙类的分支系统图

杨钟健教授是忠诚的爱国主义者、杰出的科学家和教育家,中国古脊椎动物学的奠基人和中国科学院古脊椎动物与古人类研究所的创始人。他一生的学术活动都是与我国古脊椎动物学的发展历史紧密联系在一起的。早年(1925—1935)他主要从事我国北方新生代地质及哺乳动物化石的研究。曾领导周口店的发掘和研究工作。在1935—1948年以较大精力投入爬行动物化石的研究,包括对新疆和山西的三叠纪爬行类,四川、新疆、内蒙古侏罗纪和白垩纪的恐龙化石的研究,特别是对禄丰蜥龙动物群的发现和研究做出了杰出的贡献。新中国成立以后,他的研究领域几乎涉及爬行动物的各个门类:包括蜥臀类和鸟臀类恐龙,水生爬行类和翼龙类。他的研究成果,填补了我国古爬行动物在大的门类、化石层位和地区上的空白,向世界证实了我国是世界上爬行动物化石最为丰富、种类多样的重要地区。

胡承志研究员是我国老一辈自学成才的古生物学家。1917年生,14岁(1931年)时进入地质调查所新生代rCf究室工作。其后的10年间,曾先后在步达生(Davidson Black)和魏敦瑞(Franz Weidenreich)工作室从事技术工作,修理“北京人”和山顶洞人化石及制作模型。这些精心制作的模型在“北京人”和山顶洞人化石丢失于第二次世界大战的战火后,具有了非常重要的科学价值,成为后来人类学研究唯一可对比的实物资料。1951年秋进入中国地质博物馆(原地质陈列馆)工作,直至退休。自1957年开始投入新建成的地质博物馆的筹备工作,在贵州兴义,云南禄丰、路南,山东诸城等地开展了大规模的野外采集工作,所采化石中包括胡氏贵州龙、小禄丰兽、大可翼齿兽、路南滇雷兽、沙河短齿貘、云南德氏貘、原蹄齿犀、小巨犀,云南原始猪齿兽、最后始爪兽、进步始跑犀等的模式标本。此外,他还从事了诸多门类的脊椎动物化石研究和整理、鉴定工作,研究了元谋直立人牙齿化石,并出版了巨型山东龙专著。在后期工作中,致力于采集和收集国内的珍稀矿物标本,以及与国外有关单位交换馆藏所缺的矿物标本。在他及博物馆同仁的共同努力下,使地质博物馆成为今日亚洲的第一专业展馆。胡承志先生在脊椎动物化石和岩矿方面的业绩,功不可没。
胡氏贵州龙的形态
胡氏贵州龙为大小适中的肿肋龙,其头骨的颞颥孔比鼻孔小,头骨呈三角形,最大宽度在眼眶处,嘴部小而尖,底部微呈收缩状。牙齿为同一式。脊椎数大约为:颈20、脊椎20、荐椎3或4、尾椎37以上,脊椎的背棘低、颈肋发育很好。背肋呈肿粗现象。荐椎末彼此骨连。颈相当长。肱骨与股骨的长短大致相等,但后者较细。尺骨短而宽。胫骨与腓骨比较短。前肢趾数为3、4、4、3,后肢趾数为2、3、3、4、2。贵州龙以粗壮的肱骨、宽广的尺骨、略呈多指形的前肢区别于肿肋科的其他属。

贵州龙(成年个体背视)
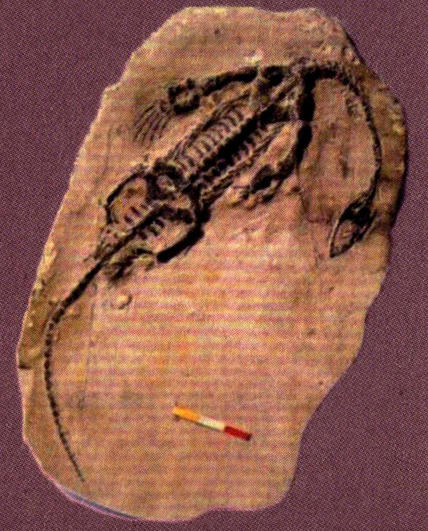
贵州龙《成年个体腹视)
贵州龙(幼年个体背视)

贵州龙(幼年个体腹视)

胡氏贵州龙复原图
胡氏贵州龙的生殖方式
胡氏贵州龙的生殖方式为卵胎生。胡氏贵州龙的肢体结构比较适合水生运动,其肘关节高度简化,而且尺骨的鹰嘴突没有骨化,若要爬上岸产卵,会是件很费劲的事;此外,同一类型的沉积物中发现了不同发展阶段的标本也支持卵胎生这一假设,但是没有直接证据证明。程延年等(2004)报道了两只怀孕的胡氏贵州龙个体。其胚胎分别位于腹中的两侧,说明雌性的贵州龙和现在的蜥蜴一样,体内存在一对输卵管。胡氏贵州龙的荐椎与骨盆之间的结构和鱼龙及沧龙中所见一样,荐肋不与荐椎结合,也不与远端髂骨结合,这样骨盆和荐骨间就允许存在相对运动,这一结构与水生性相符合。另一方面,它也允许腰带改变结构,最大化产道孔间,以加强分娩,从而说明胡氏贵州龙可能是卵胎生的。特别令人感叹的是,有两块贵州龙化石标本,记录了2.3亿年前悲壮的一瞬间,由于胚胎在母体内的体位异常(头部朝向后方),专家推断这位母亲因难产而死,随后被海底沉积物所掩埋,经过亿万年地质变化后形成了今天的化石,并为世人揭开了始鳍龙类的生殖秘密。这一重要化石的发现说明爬行动物进入海洋后不久就发展出全新的生殖方式。

怀孕的贵州龙(台中自然科学博物馆供稿)
胡氏贵州龙的双型现象
双型是指一个物种雌雄个体之间的不同形态。胡氏贵州龙为雌雄异体。雌性成年贵州龙腹中胚胎的发现,为我们辨别贵州龙的雌雄个体提供了证据:雌性成年个体较小,肱骨相对较细而平滑,骨干两端扩展不明显,肱骨稍长于骰骨,二者之比为1.05~1.16;雄性成年个体较大,肱骨相对较粗而表面粗燥,骨干两端扩张明显,肱骨长于骰骨,二者之比例为1.21~1.33。

类型多样,保存形态各异的胡氏贵州龙化石标本
胡氏贵州龙的生活习性和运动方式
胡氏贵州龙作为早期回归海洋的爬行动物,身体因适应水生生活而发生改变,尺骨与腓骨变得粗短,趾骨细长,为能游泳的典型特征。然而和鱼类动物不同,贵州龙四肢原始,未特化成鳍状,身体横宽、神经棘低、颈长、尾长,这说明它还没完全适应海洋生活,可能与现在的海狮相似,通常在近岸浅海域生活。
胡氏贵州龙的吻部小而尖,牙齿成锥状,为同一式齿形,说明它是一种肉食性动物,以鱼虾为食。
胡氏贵州龙幼年个体的前肢比后肢短,可是在以后的生长过程中,相对于身体有明显的前肢生长速度更大,而后肢则是等速或轻微的滞后。因此胡氏贵州龙成年个体的前肢比后肢更长,而且更强壮。尺骨加宽,而它的尾部神经棘较低,前部尾肋长,尾基部呈背腹向扁平;研究者推测它依靠桨状前肢的对称划动身体前进,前肢是它的运动器官。事实上也许胡氏贵州龙的运动并不这么简单,像海狮和蛇颈龙就可能混合使用两种运动方式,当前肢对称地划动时,身体后肢作背腹向运动,以产生更高的运动速度。
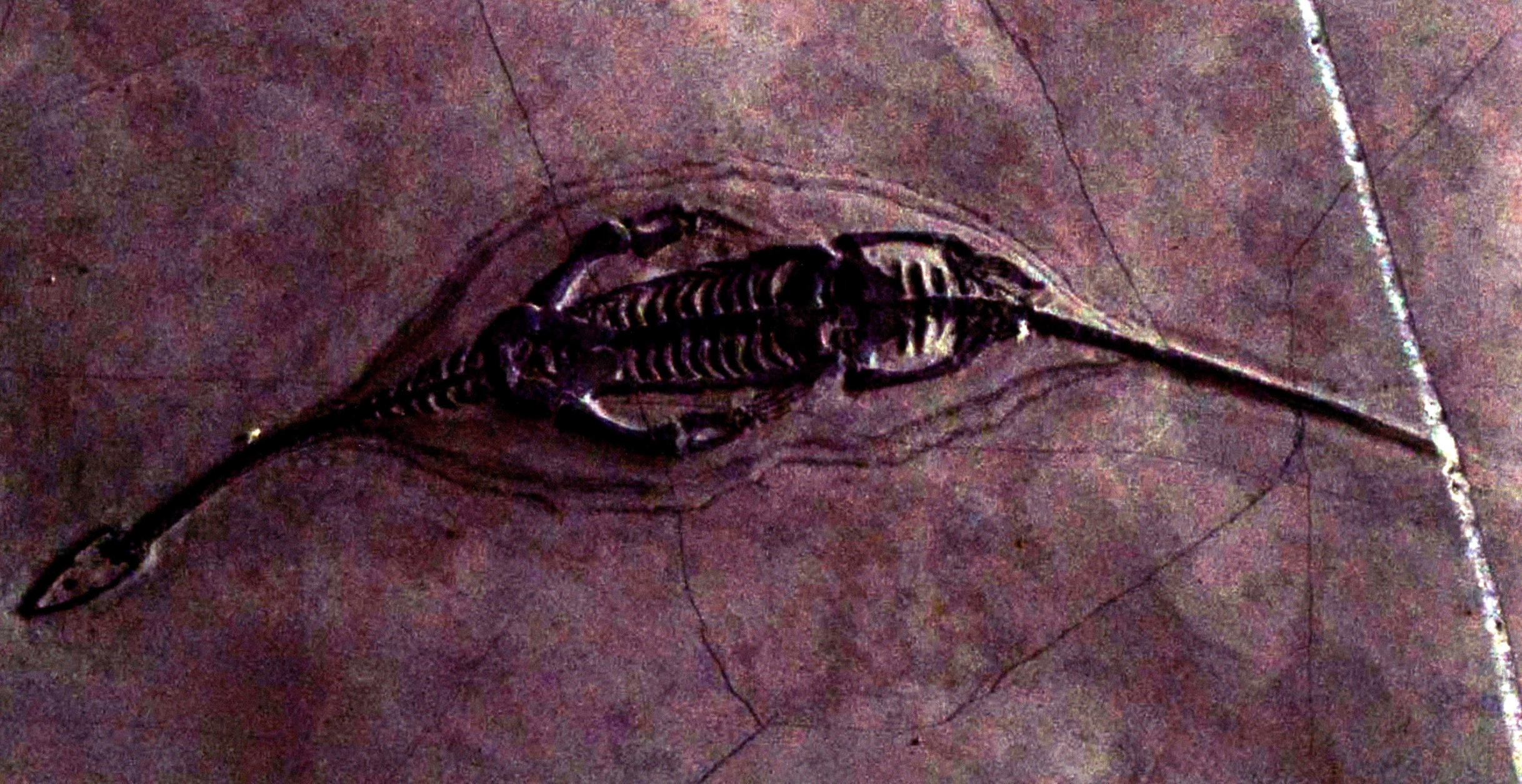
龙体长42厘米(背视)

体长28厘米、26厘米
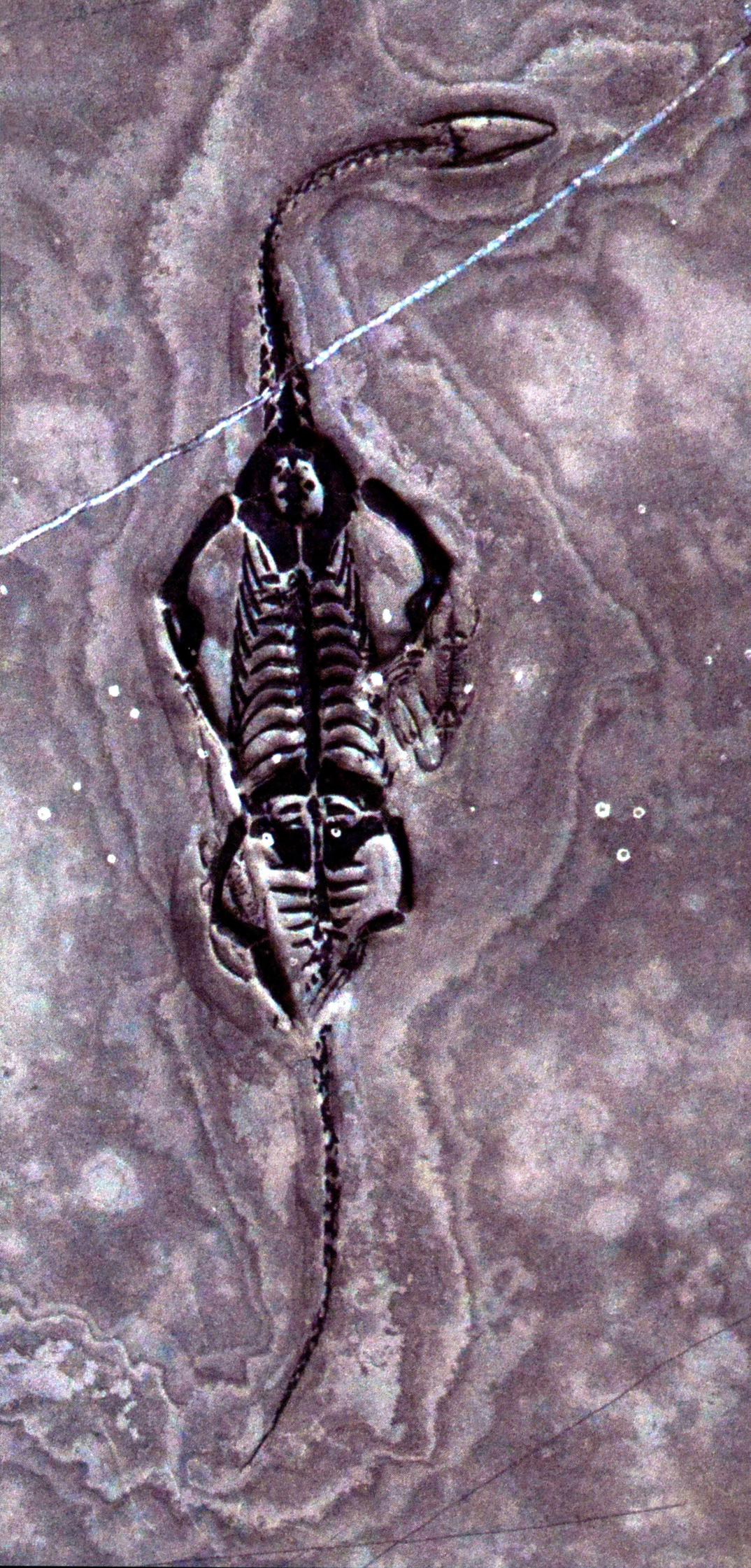
大龙长32厘米、小龙长5厘米

大龙长33厘米、小龙长约15厘米

怀孕的胡氏贵州龙(长32厘米)

怀孕的胡氏贵州龙(局部)
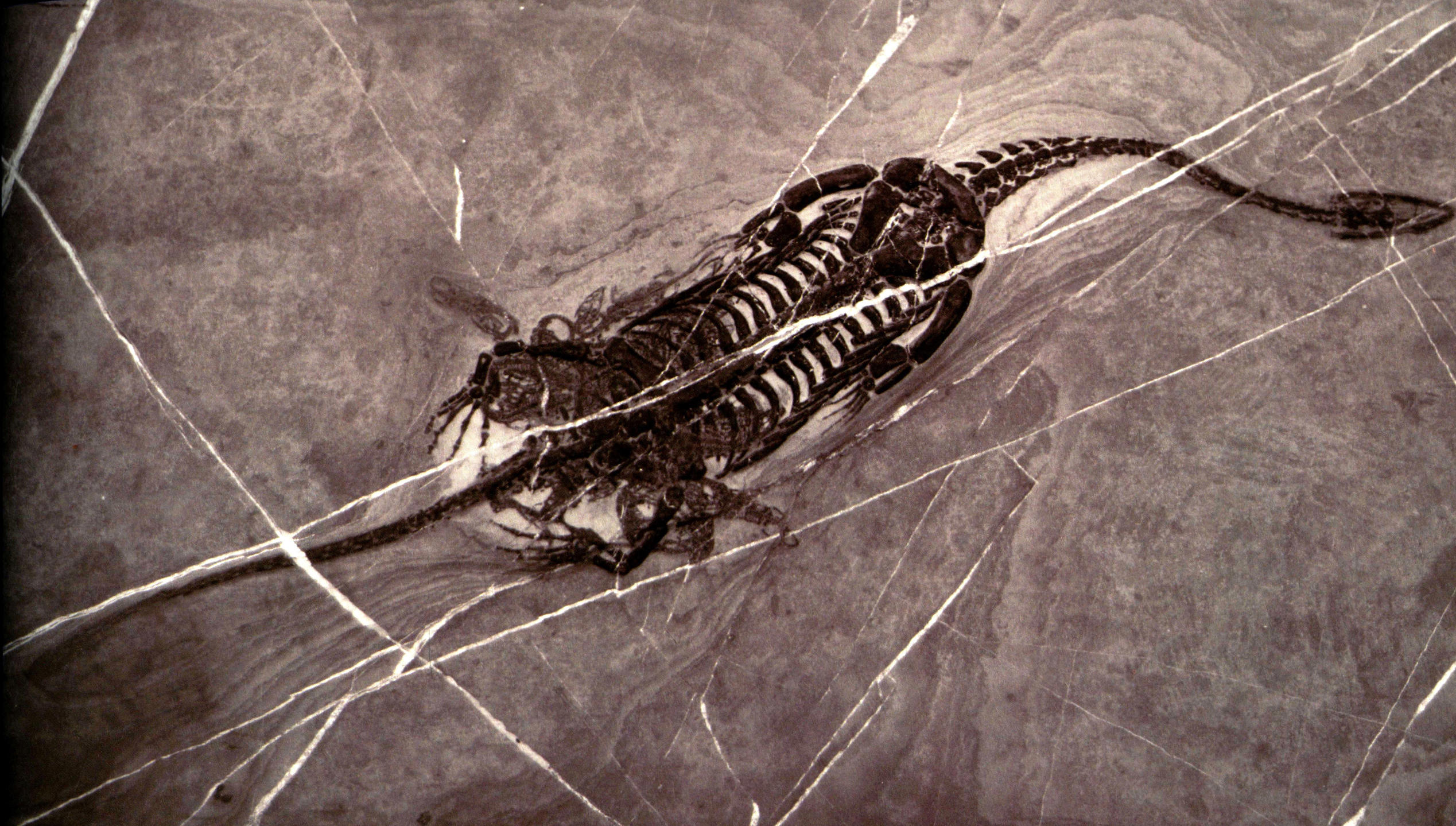
怀孕的胡氏贵州龙
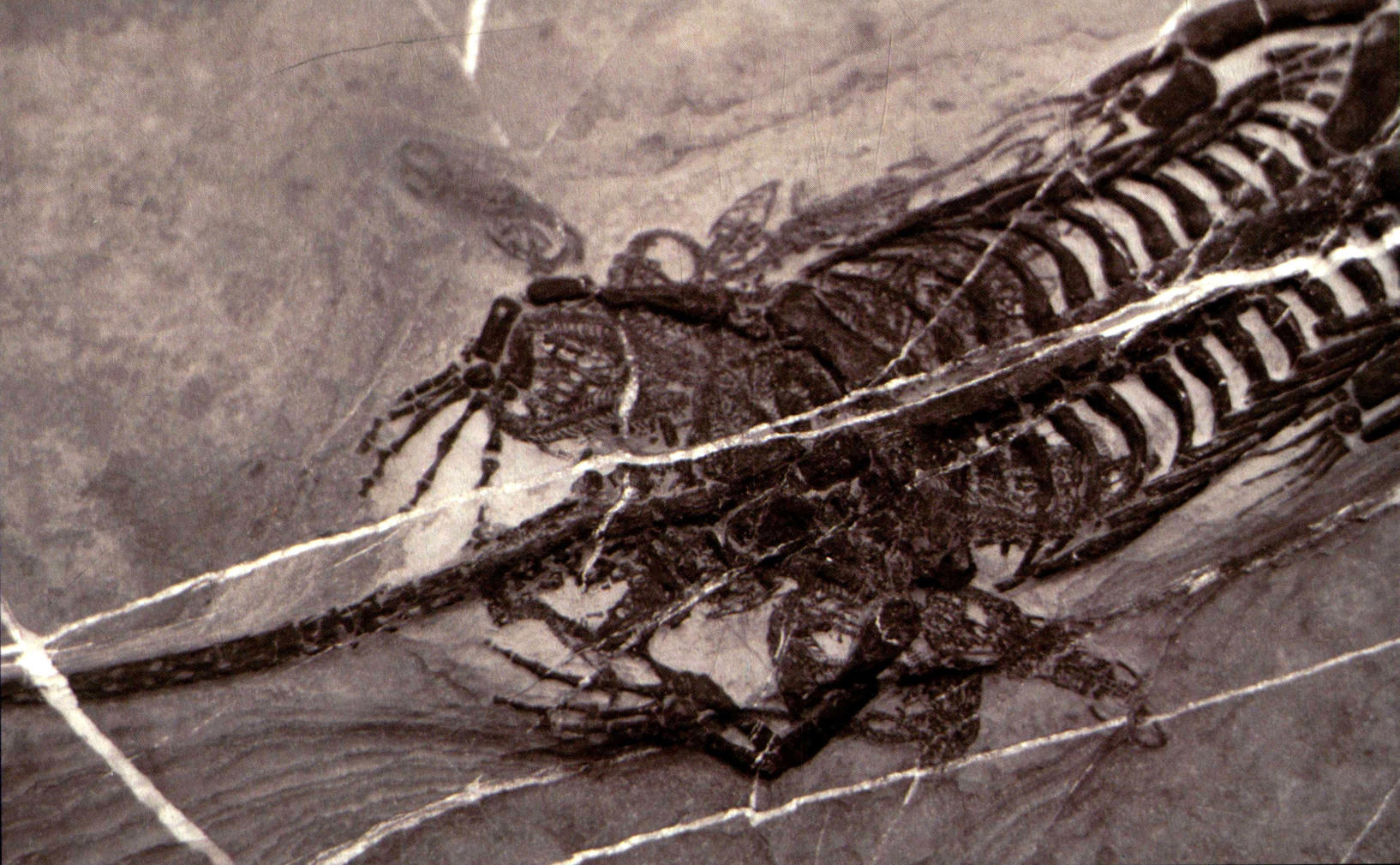
怀孕的胡氏贵州龙(局部)

体长47厘米

体长37厘米

规格22厘米×35厘米

石板规格73厘米×105厘米
细齿乌蒙龙
2008年发现于贵州省盘县新民乡羊圈村附近中三叠世早期(安尼期)关岭组二段中上部。
细齿乌蒙龙是始鳍龙类中最原始的成员之一,与肿肋龙相似,但个体比较大,体长多为1~2米,头部较小(曾被当地人称为小头龙),下颌伸长,布满了细密的垂直定向排列的牙齿。

长170厘米
二、幻龙类
Nothosaurids
幻龙类是小型到中型大小的基干始鳍龙。它的体型细长,头骨小而扁平,颈部和四肢较长;眼孔小于颞孔。长的颌骨边缘有为数众多尖利的牙齿。与肿肋龙的头骨比较起来,它的鼻孔位置更靠后,上颞孔更大,而眼孔相对较小。长的颌骨边缘有为数众多的尖利牙齿,当幻龙类在水中游泳时,它长而灵活的颈部可以左右摆动,捕食鱼类。幻龙类最早出现于早三叠世晚期(奥伦尼克期),繁盛于中三叠世,晚三叠世消失。
贵州已描述的幻龙类化石有3个属7个种,即:杨氏幻龙(Nothosaurusps;(=shingyisauyas unexpectus),n.yongi.),小吻幻龙(N.rostellatus)、羊圈幻龙(N.yangjuaneusis),兴义鸥龙(Lariosaurusing-yiensis),红果鸥龙(L.houngguoesis),邓氏三桥龙(Sanchaosaurus dengi)以及幻龙未定种(Nothosaurus)=(shingyisaurusunexpeclus)。

幻龙复原图
中国第一块幻龙化石
1965年,杨钟健教授记述了采自贵州顶效,与胡氏贵州龙同一层位,同一地点幻龙类新材料。化石是一块受挤压的不完整头骨和5个前部颈椎,它被定名为意外兴义龙(Sningyisaurus unexpeden),归入鳍龙类目扁鼻龙科。后人在1988年对化石进行了重新研究,发现它具有一系列幻龙属的特征,如尖锐微弯曲的牙齿,后置的松果孔,额骨—顶骨骨缝在上颞孔前缘之后等。因化石保存过于残缺,很难找到这一化石有别于幻龙其他种的鉴定特征,原定属种名称“意外兴义龙”被废弃,化石作为幻龙属的未定种处理,这是幻龙属在中国的第一个代表。

中国的第一块幻龙类化石,不完整头骨背面(左)和腹面(右)
杨氏幻龙
Nothosaurus youngi
杨氏幻龙产于贵州省兴义市乌沙镇、顶效镇等地法郎组竹竿坡段下部,其时代为中三叠晚期(拉丁期)。
杨氏幻龙为幻龙属的一个小型种,吻部适度发育,吻端钝圆,后部有明显的收缩,具5个匍匐状的前颌骨獠齿和4个弯曲向上的下颌缝合部獠齿,细条状态的轭骨前端未伸达眼眶边缘,上颌骨和眶后骨在轭骨之后相接,眶后弓窄,外翼骨形成明显的腹问凸缘,下颌骨凸缘,下颌骨具清晰的冠状突,夹板骨前端进入下颌骨缝合部,腕骨4块。

杨氏幻龙(长156厘米)
杨氏幻龙(体长125厘米)
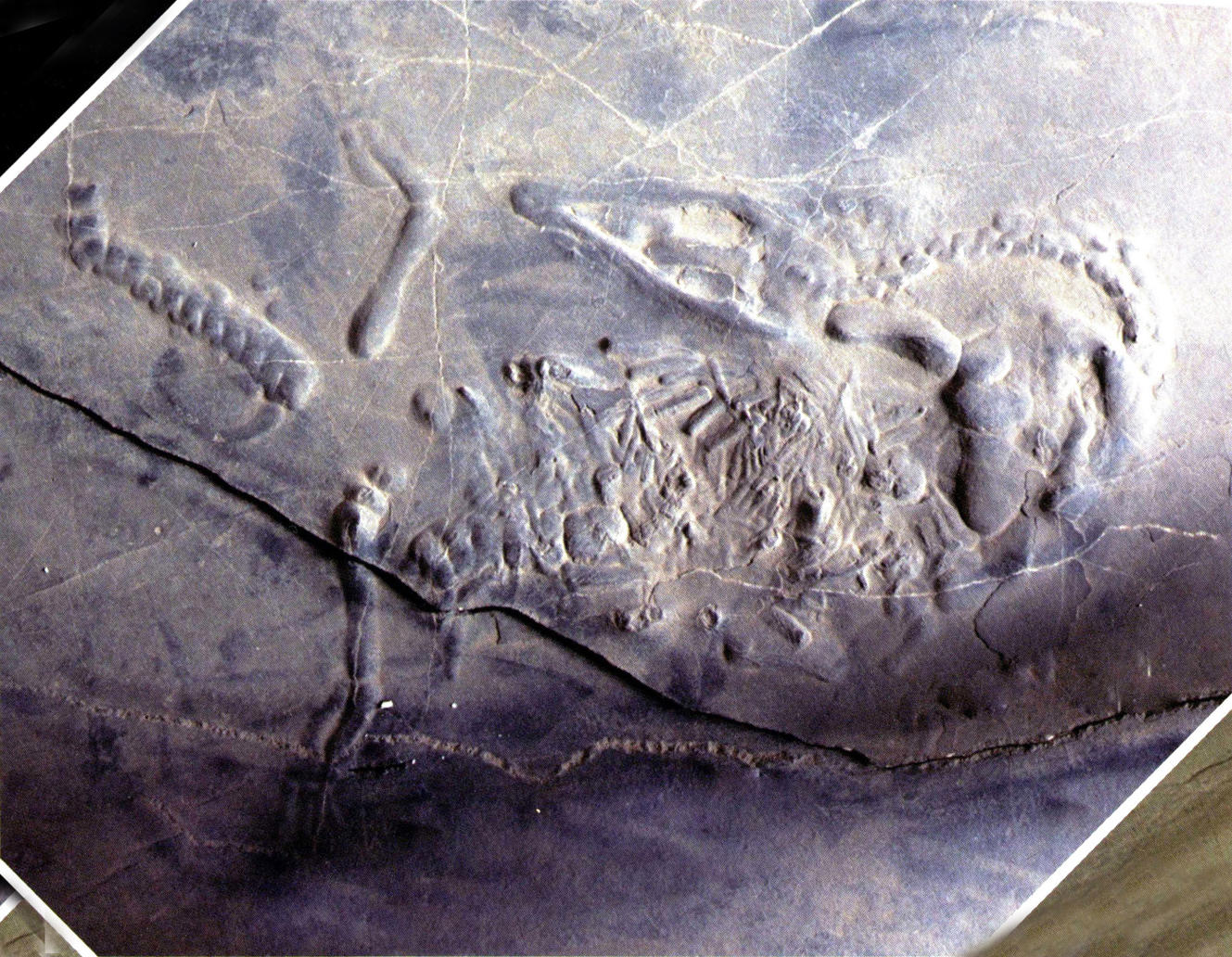
杨氏幻龙(长89厘米)
小吻幻龙与羊圈幻龙
Nofhosaurub rosfellatub and Nofhosaurus yangtuansis
小吻幻龙与羊圈幻龙均产于贵州省盘县新民乡羊圈村附近关岭组第二段的中上部,地质时代为中三叠世早期(安尼期)p1五期。
小吻幻龙头骨中线长21~32厘米,据不完全的骨架推测,其身体全长可达250厘米。吻部短小,长度略大于宽度,前端钝圆,基部收缩,具4个前颌骨獠齿,其后的是第五个明显较小的前颌齿,一对上颌骨犬齿之前有6~9个小型上颌齿,眼窝较大,卵圆形位置相对靠前,内鼻骨的后援由腭骨构成,具内鼻槽,外翼骨形成腹向凸缘,下颌骨具冠状突,背椎神经棘较低,背肋肩部明显加厚。
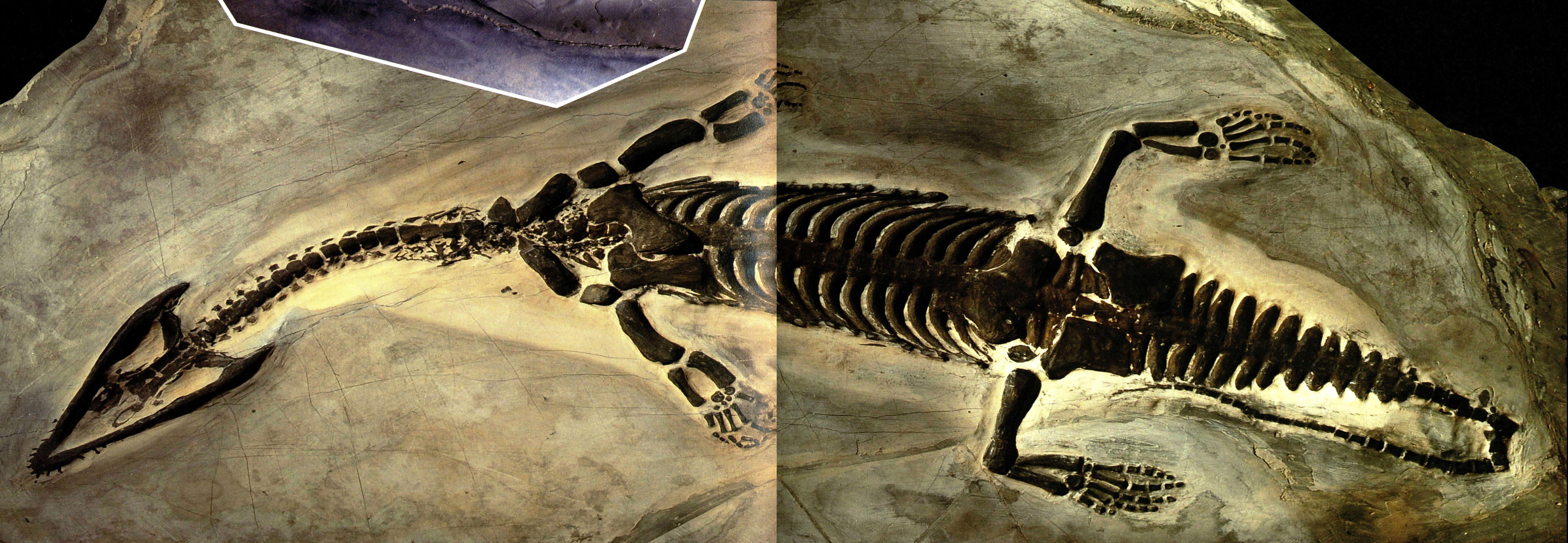
小吻幻龙
鸥龙
Lariosaurus
鸥龙与幻龙十分相似,其区别在于头后骨和腰带。鸥龙的间锁骨三角形,无后部的中央骨干。荐助5节,远端变细,彼此聚拢。腕骨5块,肱骨短。
鸥龙属现已记录两个种,即兴义鸥龙(Lariosaurus xingyiensis)和红果鸥龙(L.Houngguoensis)前者产于兴义顶效乌沙等地的中三叠世晚期法郎组竹竿坡段的下部,后者产于贵州省盘县新民乡羊圈村附近中三叠世早期关岭组第二段的中上部。
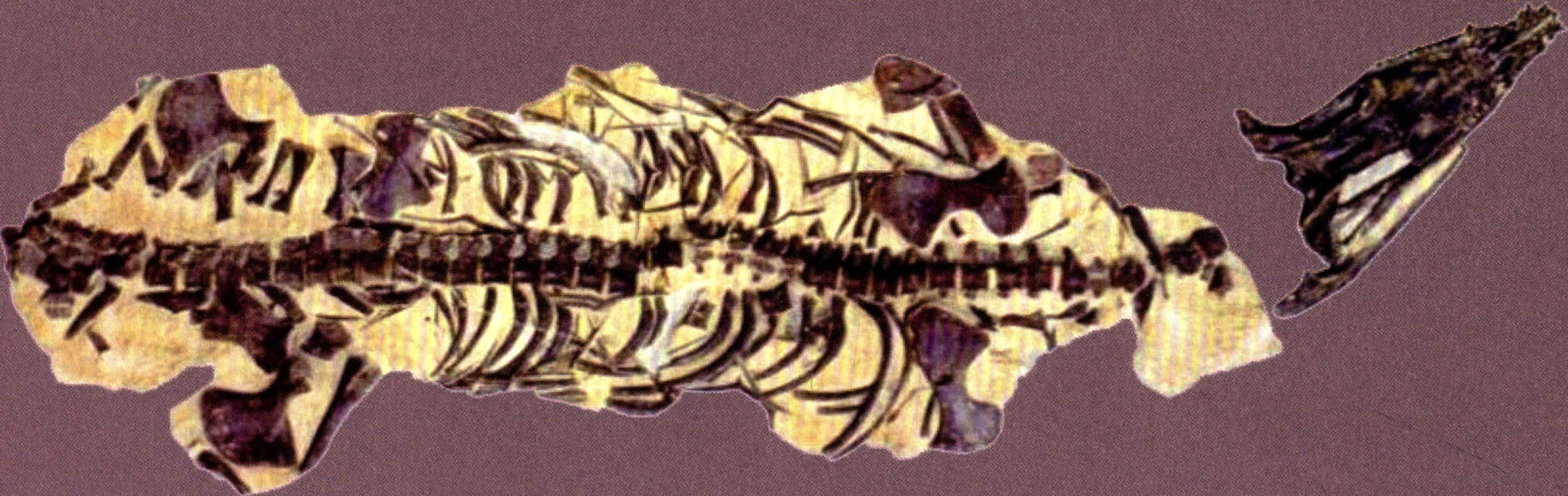
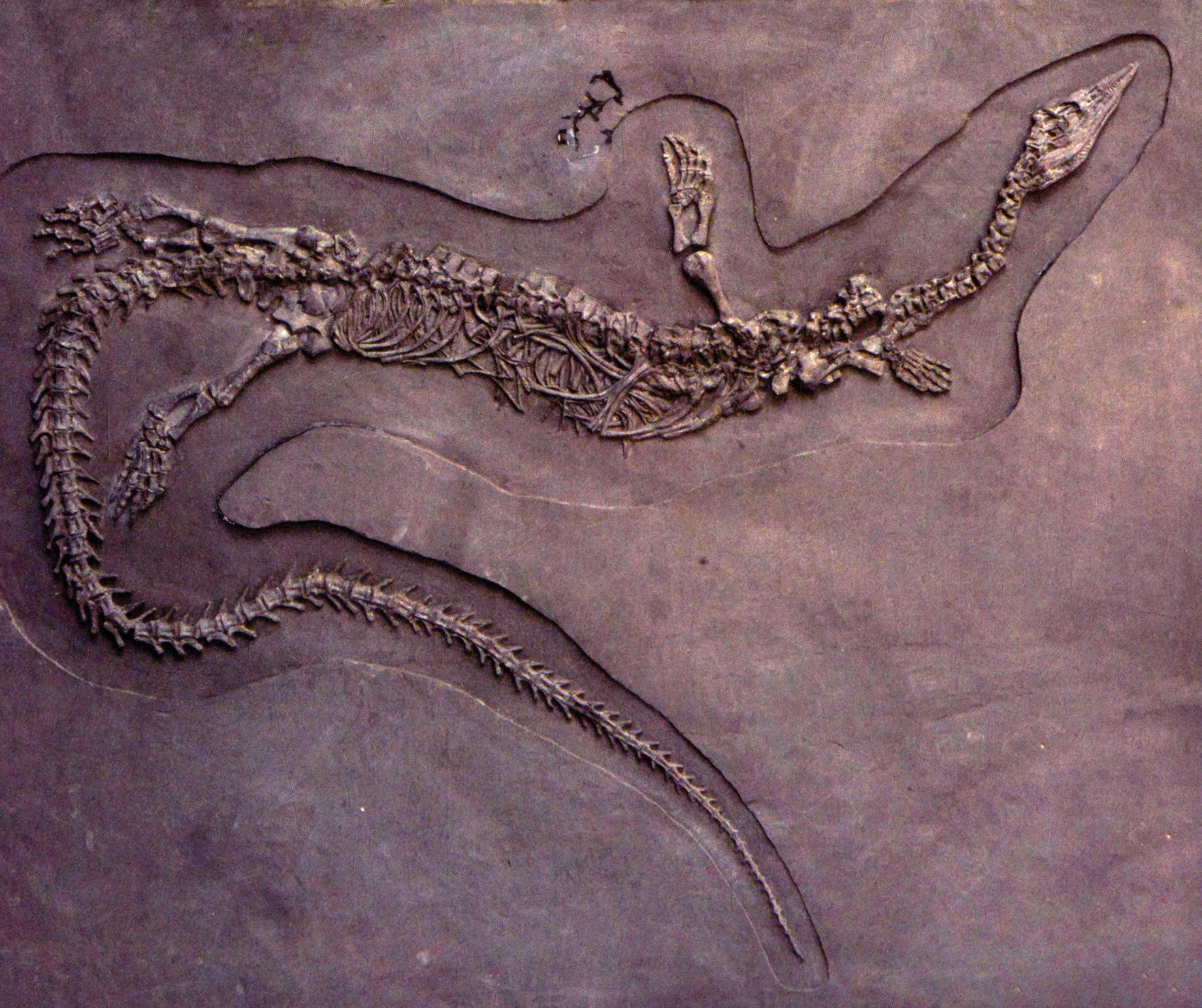
兴义鸥龙的正型标本是一个呈“S”形弯曲的骨架,清楚地显示了动物在面临死亡时的极力挣扎的痛苦状态。兴义鸥龙体长不足2米,头骨细长,外鼻孔之前吻的两侧,有明显的收缩,最前端有5个大而尖锐的、表面具清晰纵向条纹的前上颌骨齿,这牙齿足以证明它们是浅海域凶猛的捕食者。兴义鸥龙的眼眶宽大,而上颞孔长度是眼眶长度的2倍,这表明有大的下颌收缩肌附着在上颞孔内。当它在捕食时,肌肉的收缩可以产生强大的力量关闭上下颌。尖锐的牙齿可以刺穿鱼类,或许还有其他软体动物的身体。

兴义鸥龙(长200厘米)

红果鸥龙头骨

鸥龙生态复原图
三、纯信龙类
Pistosauroids
纯信龙是个较少出现的类群,到目前为止在中国仅有零星发现,但纯信龙在海生爬行动物进化上占有重要的位置——它是幻龙类的近亲,是繁盛于侏罗纪、白垩纪蛇颈龙的祖先类型。纯信龙的分类位置在不同学者的不同分类系统中虽然有所差异,但研究者都认为它是自幻龙向蛇颈龙类进化的不可或缺的中间环节。纯信龙类头骨的许多特征类似于幻龙类,细长的吻部前端,向前伸出的大的前颌骨齿,但它腭面结构比幻龙原始,并未被骨片完全封闭。
宋氏清镇龙
Chinchenia sungi
产于贵州清镇关岭组中,其时代为中三叠世早期(安尼期),是贵州最早发现的纯信龙分子,但化石十分破碎,难以说明其特征。
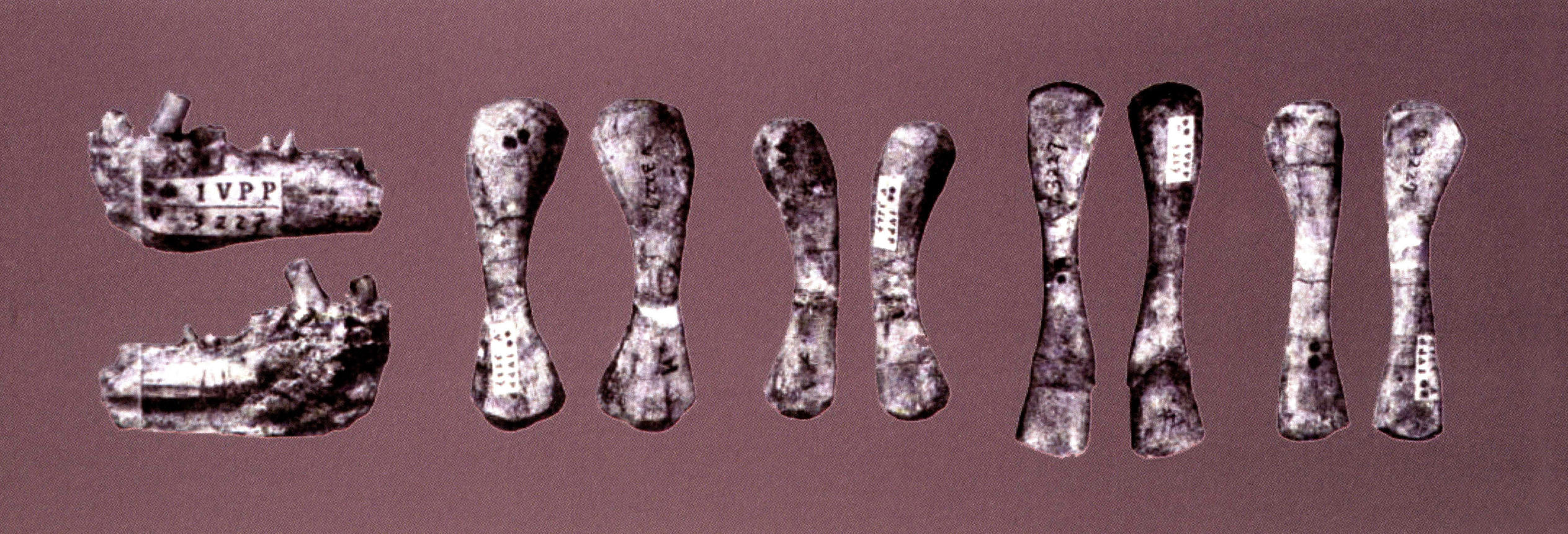
宋氏清镇龙的不完整下颌和头后骨骼材料
李氏云贵龙
Yunguisaurus liae
李氏云贵龙采自贵州兴义法郎组竹竿坡段,它是中国记述的第一个最完整的骨架,也是世界上最大最完整的纯信龙化石。化石保存长度1.7米,头骨的吻部细长,每侧着生7枚前上颌骨齿,松果孔的位置靠前,位于额骨和顶骨的骨缝上,顶骨形成类锐的顶骨嵴,下颌的缝合部长,下颌上具明显的冠状突。

李氏云贵龙(台中自然科学博物馆供稿)
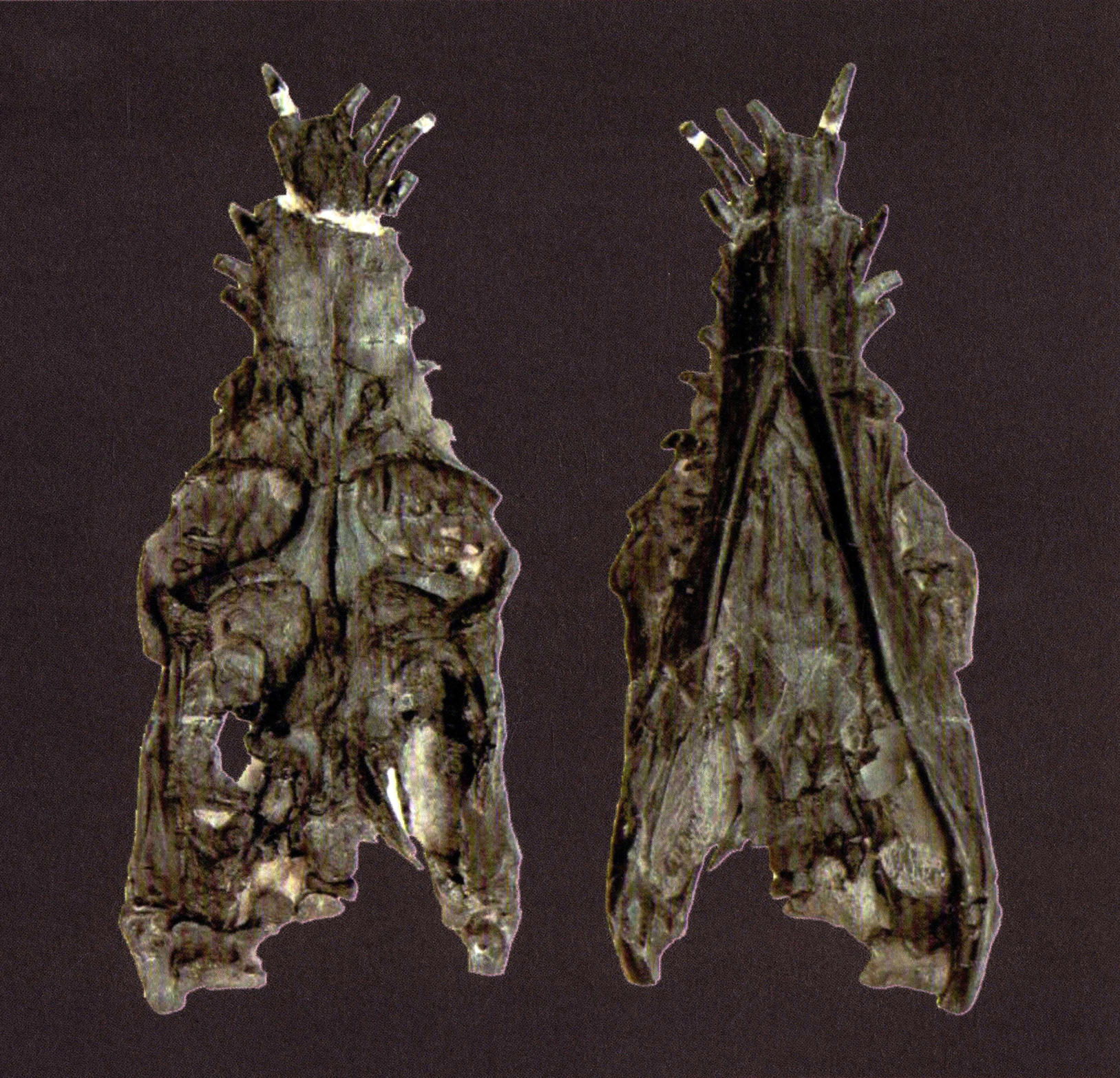
李氏云贵龙头骨(台中自然科学博物馆供稿)

云贵龙复原图
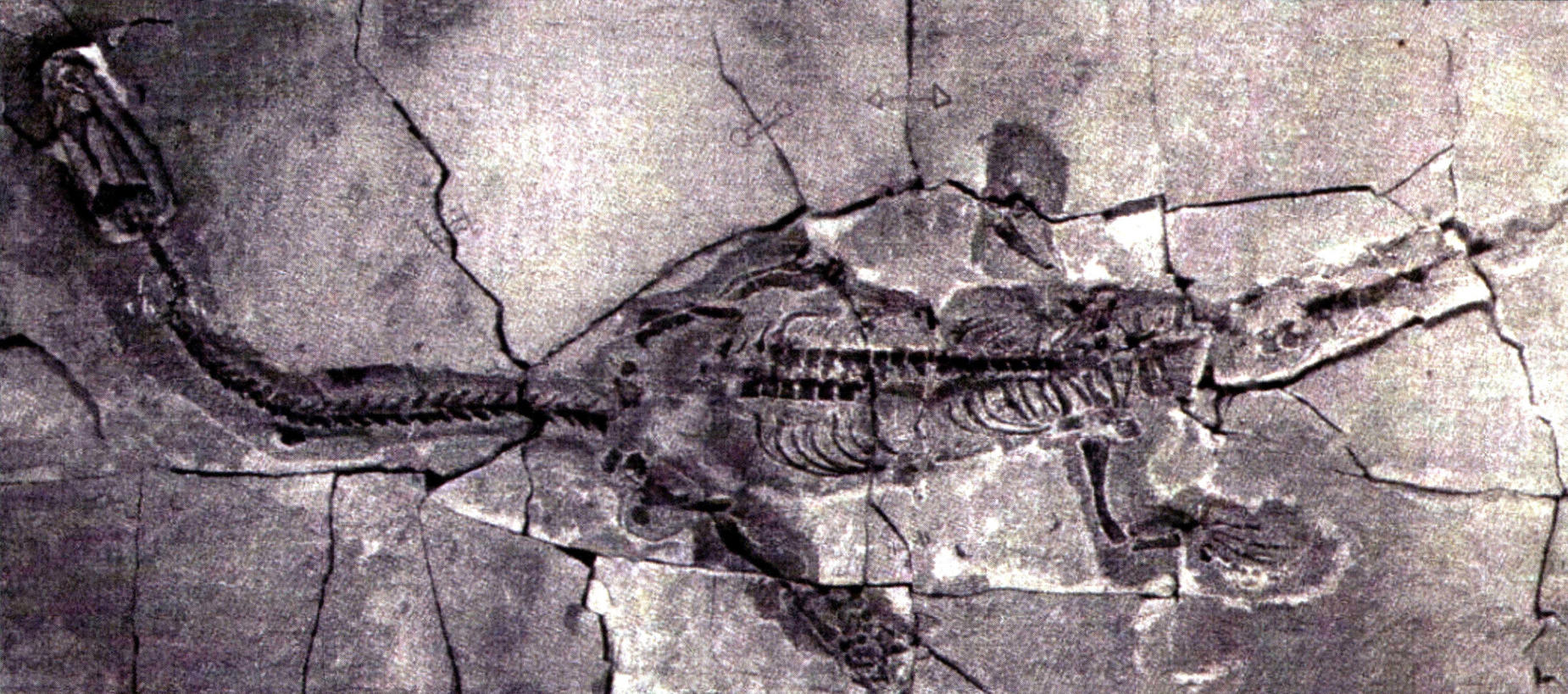
短吻王氏龙正形标本骨架
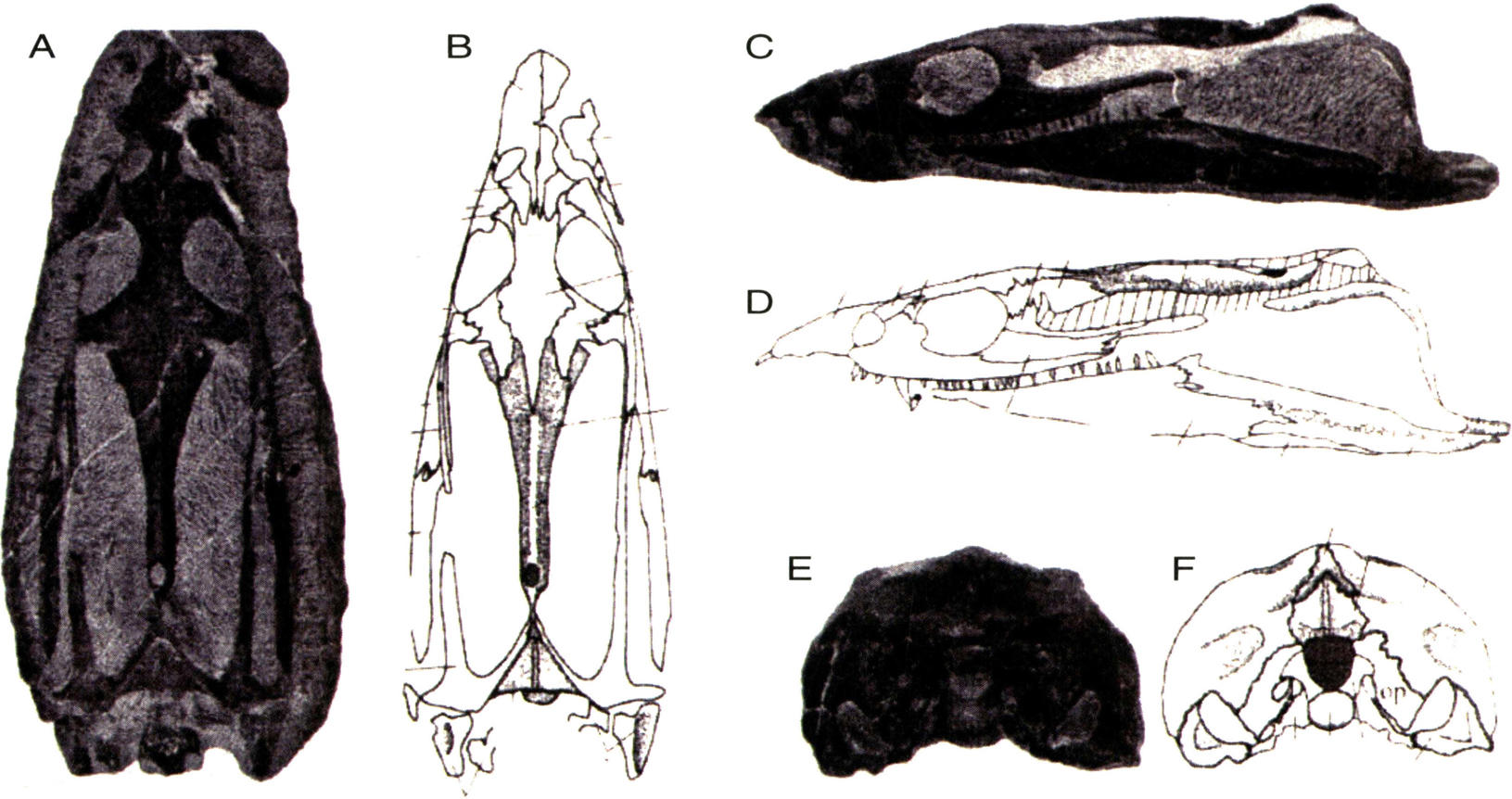
短吻王氏龙头骨(A-B背视,C-D侧视,E-F后视)

幻龙(大龙长192厘米,小龙长70厘米)

幻龙复原图

幻龙(长150厘米)

幻龙(此件化石很特殊,因地壳作用石板变成扭曲形状,但龙化石保存十分完整,体长140厘米)

幻龙(体长149厘米)

幻龙(长280厘米)

幻龙(长95厘米)

幻龙(长250厘米,未修复)
幻龙正负模板,上图为副模版,下图为正模版。(长130厘米)

幻龙俯视和背视,平板化石出现正反面都同时能修出化石的十分稀少。(长166厘米)
幻龙群谱(规格:270厘米×150厘米)
四、待定类群
不能归属于肿肋龙类、幻龙类、纯信龙类的始鳍龙类,尚有以下3种。
利齿滇东龙
Diandongosaurus acutidentatus
滇东龙是小型鳍龙类(模式标本长28.5厘米),既有肿肋龙吻部两侧不收缩,眶前区域长于眶后区域,眼眶大于上颞孔等典型特征,又具有幻龙类前颌和下颌前部大齿型齿发育,上颌具1或2个大齿型齿,顶骨和额骨均愈合,顶骨平台收缩,前后关节无肿胀现象等特征。
滇东龙额骨两后外侧支的末端后延超过上颞孔的前缘,轭骨和鳞骨相交将眶后骨排除于下颞孔之外,方轭骨发育,锁骨前外侧缘具一突起,三对荐肋以及最前部尾肋的外侧均未见明显收缩,这些特征也多表现出肿肋龙和幻龙类的混合特征。
除此之外,滇东龙还具有一些有别于肿肋龙和幻龙类的较为独特的特征,包括前额骨和后额骨沿眼眶背缘相交,轭骨和鳞骨相交,鳞骨和方骨支伸达至方骨的关节,第3对至第4对尾肋的长度超过荐肋,后肢未端发育异常膨大的瓜趾骨等。
滇东龙与贵州龙的区别主要是头后骨骼的差异,滇东龙背长大于颈长,肋骨无肿胀现象,尺骨未显著加宽,仅有两个骨化的脘骨。
系统分析表明:滇东龙既不是肿肋龙类,也不是幻龙类,为始鳞鳍类的基干类群。化石产于云南省罗平县罗熊镇大洼子村附近中三叠世早期关岭组第二段中上部。

利齿滇东龙(长45厘米)
丁氏滇肿肋龙
Dianpachysaurus ding
产于云南省罗平县罗熊镇大洼子村附近关岭组第二段中上部,其地质时代为中三叠世安尼期pelsonian亚期。

丁氏肿肋龙骨架(正形)
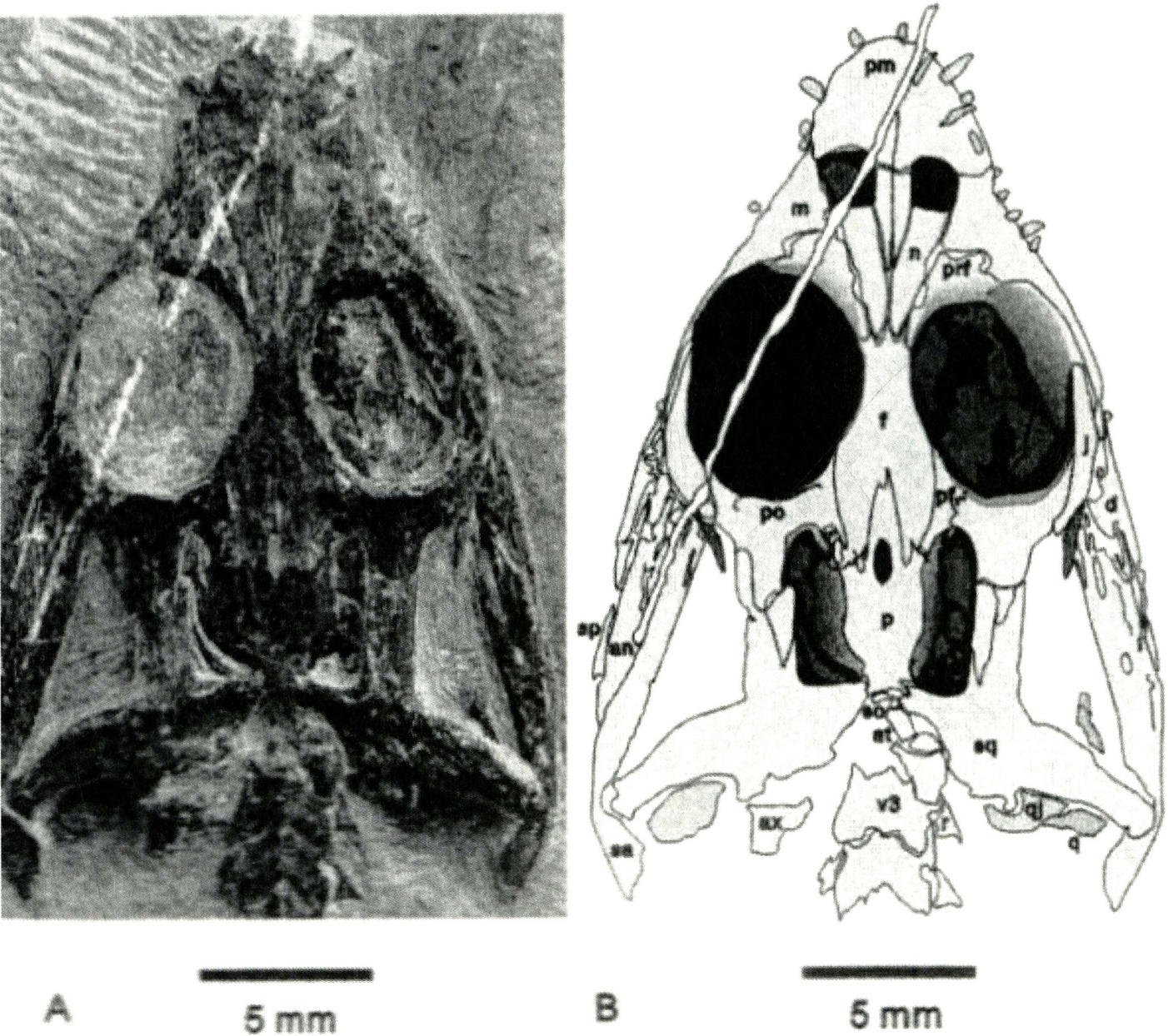
丁氏肿肋龙头骨(正形)
岔江黔西龙
Qianxisanurus chajiangensis
产于兴义市乌沙镇岔江村附近中三叠世晚期(拉丁期)法郎组竹竿坡段。

优美滤齿龙
楯齿龙
Placodonts

楯齿龙上、下颌骨及头部的颚部都生长着形似蚕豆的大垂板状牙齿,这些牙齿好像小磨盘,在强有力的下颌肌肉的带动下可以轻易地压碎软体动物的外壳。楯齿龙可以用“楯齿”对付无脊椎动物的甲壳,同时用厚重的壳抵御其他体形巨大的海生爬行动物。
楯齿龙类扁平的身体和强壮的四肢表明,他们不是快速游泳的动物,只能在浅海环境中用四足在水中划动,推动身体缓慢前行。
楯齿龙类是外形十分奇特的三叠纪海生爬行动物。根据甲壳的有无,楯齿龙类可以划分为两大支系。其中大部分种类属于有甲壳的一支,即豆齿龙类,豆齿龙的外形与龟相似,颈短、身体变得很宽而且扁平,表面有骨质骨片形成的厚重背甲,在一些种类中还发育了腹甲,腹甲通过侧壁与背甲相连,一些属种的背甲后壁面还有一块较小的“第二背甲”。单独的覆盖着尾巴前面的部分,还有些种类的整条尾巴上都覆盖着骨板;另一类是楯齿龙类,它还没有进化出豆齿龙类那种完整的甲壳,只在背部从前向后排列着一些零散的甲片,它不仅具有大量板状齿,而且口腔前面的牙齿粗壮而钝圆,并且向前突齿而出,可以同时发挥“凿子”和“钳子”的作用,将附着在岩石上的猎物揪下来。
目前发现的楯齿龙化石有4属4种:
豆齿龙类,即新铺中国豆齿龙(Sinocya modus xinpuensis)——中国第一个发现和描述的楯齿龙类化石,多板石乐甲龟龙(Psephochelys polyosteodma),康氏雕甲龟龙(Glyptopsephochelys kangi);楯齿龙类,仅有意外龟龙(Placodus inexpectatos)。

楯齿龙生态复原图
一、豆齿龙类
新铺中国豆齿龙
Sinocyamodus xinpuensis
新铺中国豆齿龙产于贵州省关岭县新铺镇一带晚三叠世法郎组瓦窑段。
中国豆齿龙是一种小型豆齿龙,其背甲较宽,由数十枚多边形的小骨板组成,身体腹面没有腹甲。背甲甲片的形状不规则,多为六边形和五边形,其中边缘的两列甲片具有明显的脊状结构,靠近颈部的部分甲片呈金字塔状。它有一条很长的尾巴,上面同样覆盖着甲片。
中国豆齿龙的四肢比较短,没有明显的适应水上生活的特征,游泳能力不强,除了在岸上休息,也只能在近岸的浅水里捕食附着在岩石上具有甲壳的动物为食。

新铺中国豆齿龙复原图

新铺中国豆齿龙(体长53厘米)

新铺中国豆齿龙(体长57厘米)

新铺中国豆齿龙和创孔海百合(体长55厘米)
多板砾甲龟龙
Psephochelys polyosteoderma
多板砾甲龟龙产于贵州省关岭县新铺镇一带晚三叠世早期(卡尼克早中期)法郎组瓦窑段。
砾甲龟龙没有腹甲,背甲非常厚重,有一个很明显的“侧壁结构”,整个背甲由多大次而枚的小甲片构成。这些小甲片以五边形和六边形为主,但形状比较规则,表面布满了细小的放射较饰。它的腹部结构十分特别,除了中部的腹肋以外,两侧还有很多不规则的长条形甲片,这种结构在已知的任何豆齿龙中都未发现。

多板砾甲龟龙

多板砾甲龟龙复原图
雕甲龟龙
Glyphoderma
雕甲龟龙产于兴义市乌沙镇一带中三叠世晚期(拉丁期)法郎组竹竿坡段。
看着很像砾甲龟类,但是雕甲龟龙的背甲结构特别精致,由400多枚表面布满了细小放射状纹饰的小甲片组成,其生活年代比砾甲龟龙大约早1000万年。

雕甲龟龙(长85厘米)

雕甲龟龙复原图
二、楯齿龙亚类
意外龟龙
Placodus inexpeclatus
意外龟龙产于六盘水市盘县新民乡羊圈村一带中三盛世早期(安尼期)关岭组第二段中部,是目前已知最早的楯齿龙化石,也是我国目前唯一发现的楯齿龙亚类的化石。

小型龟龙
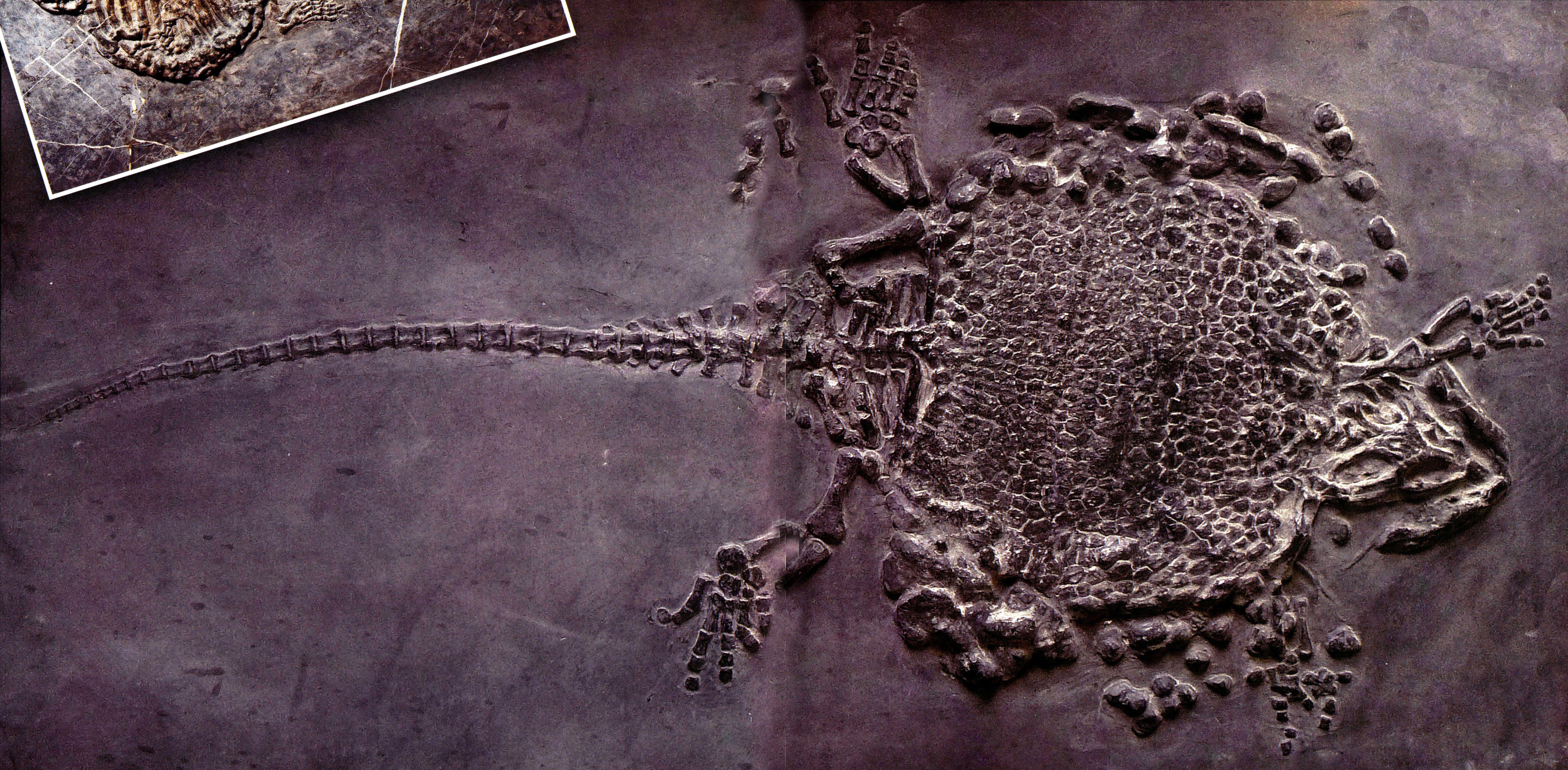
楯齿龙(长91厘米)

楯齿龙(规格36厘米×93厘米)
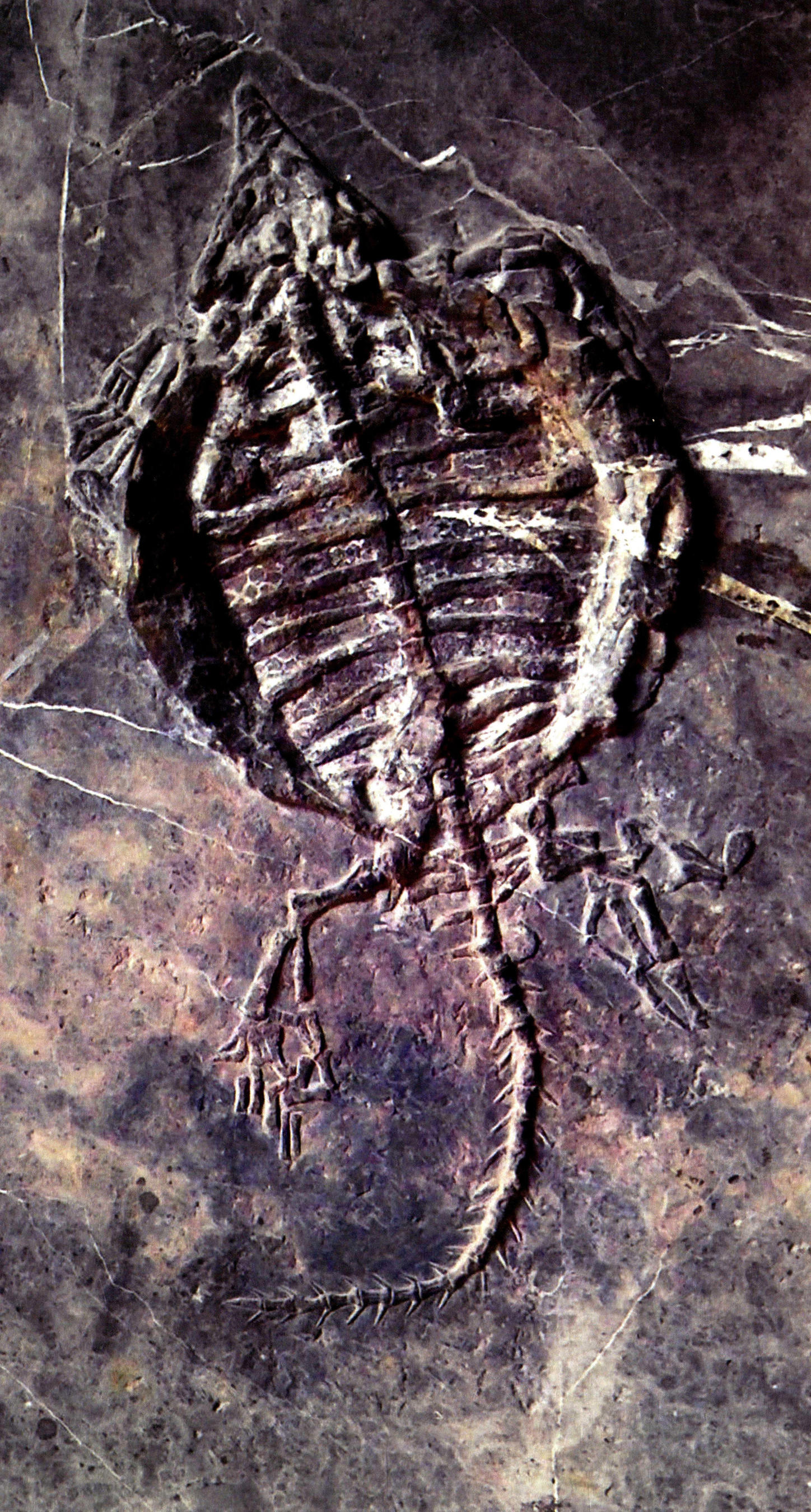
楯齿龙(长63厘米)

未定名(长70厘米)
鱼龙类
Lchtyosaurids

鱼龙是古生物学家最早(1810年)发现的中生代海生爬行动物,也是有史以来最特化的海生爬行动物,生活在中生代海洋的各个角落。它们具有优美的流线型体形,可以最大限度地减小水的阻力;鱼龙的四肢已经特化成桨状,能够像鱼类的偶鳍一样在游泳过程中保持身体平衡;作为推进器的尾部看上去像鱼类的尾巴一样呈新月形,在古脊椎动物中,就与身体的比例而言,鱼龙的眼睛是最大的,这说明鱼龙的视觉器官非常发达,可以在黑暗的水中捕食猎物。
中国第一次发现的鱼龙类
1965年,杨钟健教授依据来自贵州省仁怀县茅台镇的一零散保存的头后骨骼材料建立了混鱼类的一个新种——茅台混鱼龙(Mixosaurus maotaienois),虽然因“茅台种”的材料过于破碎,不具独有的鉴定特征,这一命名受到大多研究者的质疑,被认为是无效种,但这是中国三叠纪鱼龙的首次报道,证实了中三叠世在中国及至东亚的存在。
鱼龙化石是分布最广、延续时代最长的海生爬行动物化石,在盘县、兴义、关岭三个生物群都有产出,学界公认的有下述几种。

1965年记述的茅台混鱼龙
盘县混鱼龙
Mixosaurus panxianensis
发现于贵州省盘县中三叠统关岭组二段的盘县混鱼龙是中小型鱼龙,体长1米左右。头骨侧面可见大而圆的眼孔和尖而长的吻部。上颌上稀疏地着生16~20枚牙齿,下颌齿18枚。齿列前部为细、尖锐的锥状齿,最后部3~4枚牙齿的齿冠较粗壮且圆钝。与其他的鱼龙一样,盘县混鱼龙的颈部极短;脊椎椎体侧视的高度大于前后长度;神经棘高而窄,中部背脊的神经棘是椎体高度的2.8倍;中部尾椎也有加大的现象。

盘县混鱼龙(长88厘米)

盘县混鱼龙(长155厘米)

规格:147厘米×130厘米

盘县混鱼龙(带多条小龙,长120厘米)

盘县混鱼龙(长110厘米)

盘县混鱼龙(长100厘米)

盘县混鱼龙复原图

这条盘县混鱼龙体内孕育了12条小鱼龙,是已知胚胎数目最多的鱼龙化石标本

盘县混鱼龙(长130厘米)
磨齿新民龙
Xinminosaurus catactec
新民龙是发现于贵州省盘县三叠纪中期地层的中等大小的鱼龙。它们虽然不是最古老的鱼龙类群,却保留了很多比较原始的特征,如较短的吻部以及相对保守,更近似于陆地动物的指(趾)节形态等。最为奇特的是,新民龙的口腔后面生长着两排宽大而粗壮的豆状牙齿,这可能与它们的特殊食性有关。

新民龙
周氏黔鱼龙与兴义黔鱼龙
Qianichthyosaumus zhoui and Q.xingyiensis
周氏黔鱼龙产于贵州省关岭县新铺镇附近晚三叠世早期法郎组瓦窑段。
兴义黔鱼龙产于兴义市乌沙镇附近中三叠世晚期法郎组竹竿坡段。
黔鱼龙体长多为1~3米,吻部短而尖,脑颅相对较大,因而整个头骨看上去与鸟类的头骨轮廓近似。一般来说,三叠纪的鱼龙与侏罗纪和白垩纪的鱼龙在体形方面有明显差异,三叠纪鱼龙尾椎后部向下弯曲的角度较小,黔鱼龙尾椎弯曲的角度恰好介于两者之间,其背部的弯曲弧度也远远大于三叠纪同期的鱼龙。这些特征表明,黔鱼龙很可能是一种过渡型的鱼龙。

周氏黔鱼龙(220厘米)

有多条幼体的周氏黔鱼龙(长210厘米)

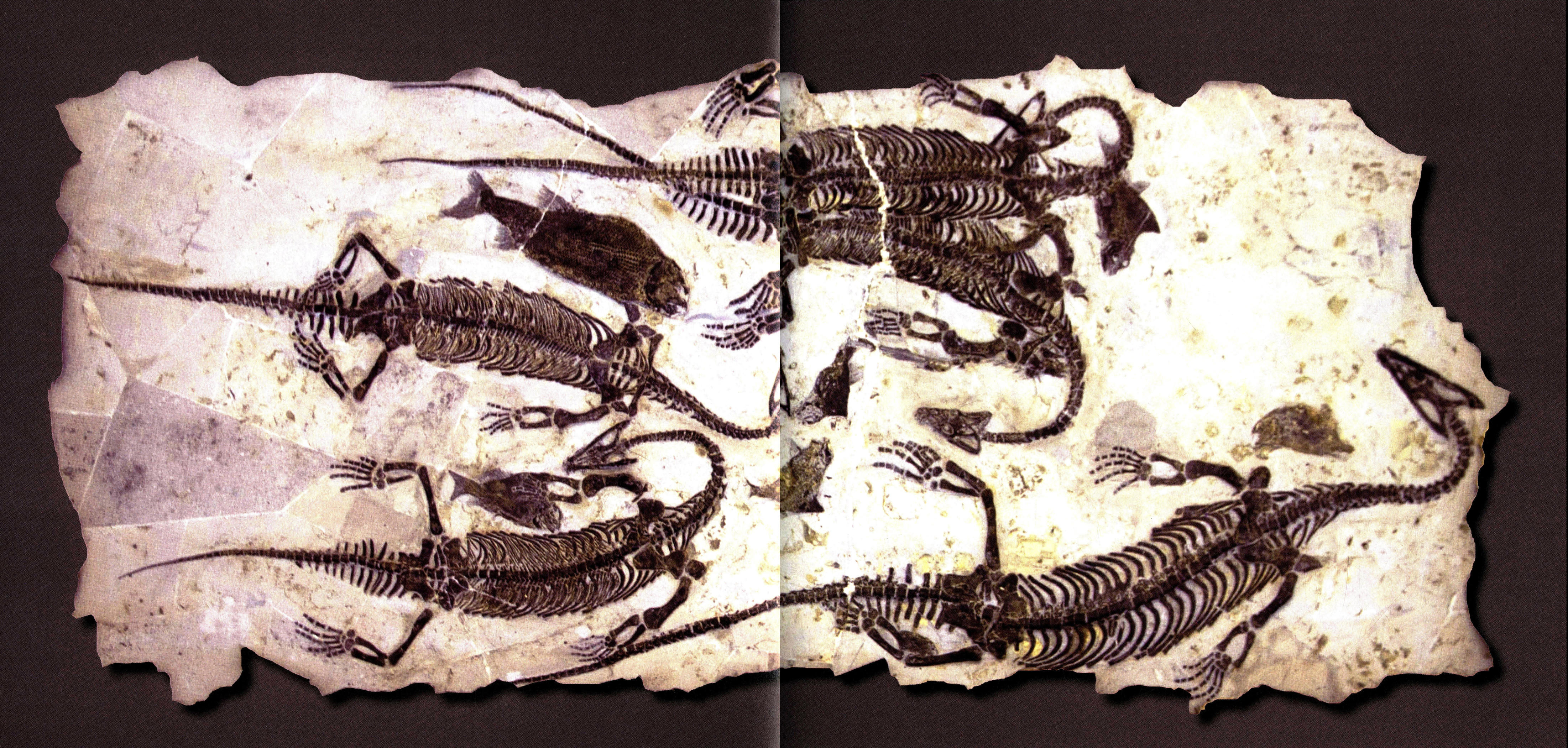
黔鱼龙和多条幼体黔鱼龙与创孔海百合在一起(龙长203厘米)。鱼龙是“胎生”还是“同类相食”的争论一直没有间断。早在19世纪中期(1846年),古生物学家就在英国西部地区发现了体内保存有幼体的鱼龙化石,并提出鱼龙胎生的假说。此后,英国和德国的侏罗纪地层中陆续又发现了不少这样的标本。但它们到底证明了什么,是胎生?还是同类相食?学术界就此曾发生过激烈的争论。要知道,成年个体捕食幼体的现象在现生爬行动物中并不鲜见。通过对标本的仔细分析,包括小鱼龙在大鱼龙体腔内的位置以及其骨骼的关连程度,胎生的观点逐渐占据了上风。最后,产自德国的关键标本为这一假说提供了无可辩驳的证据,化石清楚地显示了鱼龙产子的瞬间。

黔鱼龙(下图局部)

黔鱼龙与创孔海百合(规格302厘米×182厘米,龙长410厘米)

黔鱼龙(长230厘米)

黔鱼龙(长180厘米)
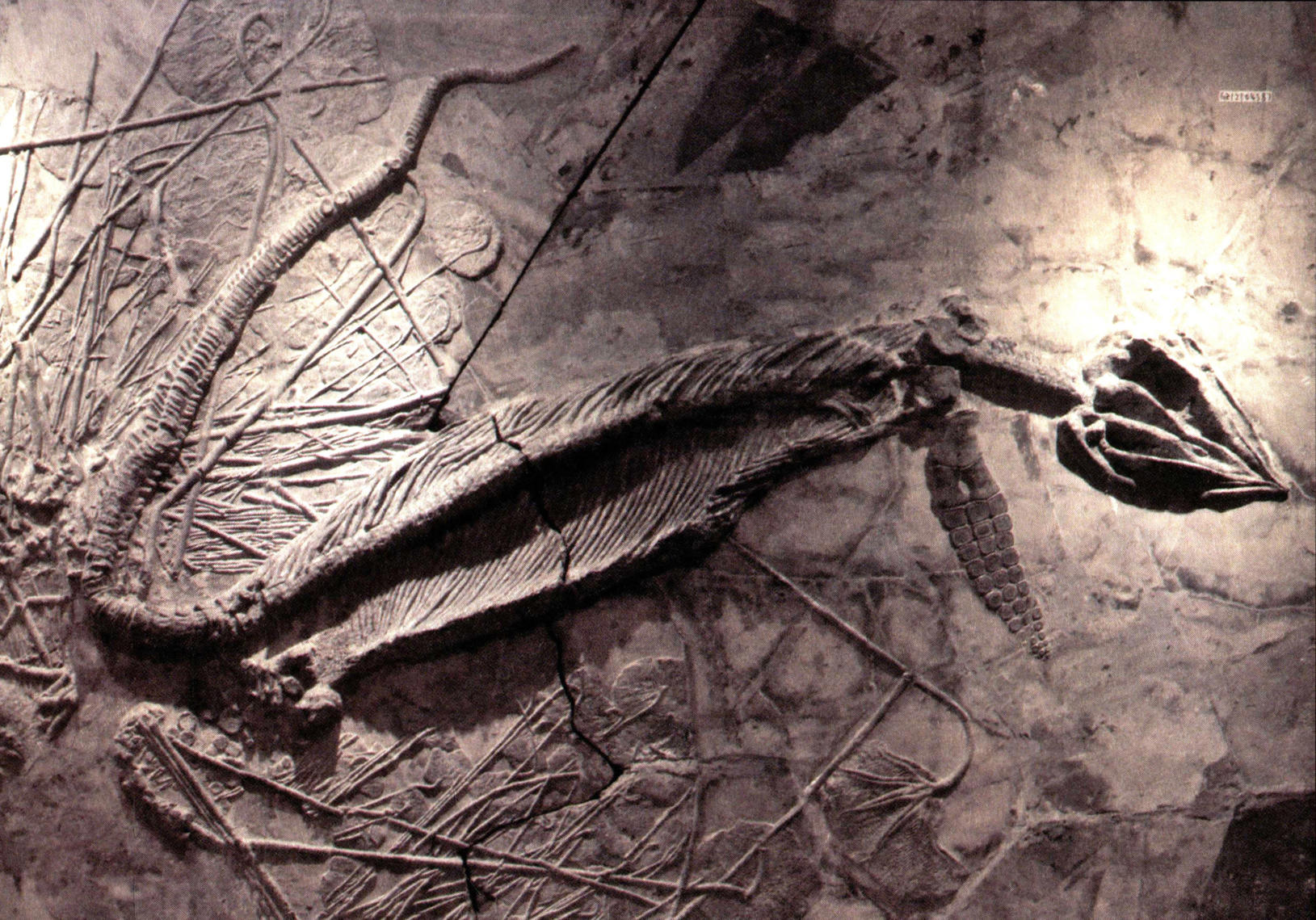
萨斯特鱼龙与海百合(长300厘米)

黔鱼龙(规格198厘米×84厘米)

周氏黔鱼龙(规格178厘米×88厘米,体长205厘米)

萨斯特鱼龙(双龙,体长分别570厘米510厘米)

萨斯特鱼龙(未定属,长430厘米)与创孔海百合
萨斯特鱼龙
Shastasaurus
萨斯特鱼龙目前仅描述一个种,产于贵州省关岭县新铺镇地区晚三叠世早期(卡尼期)法郎组瓦窑段,为一大型、长吻和窄鳍型的鱼龙类。头骨顶视为长三角形,吻长,眼眶中等大小,颞孔小于眼孔;松果孔小,为椭圆形;鼻孔小。牙齿为圆形、圆锥形。颈椎较短,荐前椎约分58~60节,椎体短,圆筒状,双凹型,前后端略扩展,中部略下凹,横突较低,肋骨较短,单头型,肋头扁平,肋骨中部有一自近端向远端延伸的浅沟。肩胛骨粗短而厚,乌喙骨较大,板状,近于半圆形;肱骨宽大,近于圆形,桡、尺骨近方圆形;腕骨3枚,近于圆形;掌骨3枚,近圆形;指骨4列,疏松排列;股骨长而厚,两端略扩展;胫、腓骨近方形;跗骨和蹠骨都近于圆形;趾骨4列,骨节为圆形,排列疏松。
最初曹泽田、罗永民(2000)将该种定为邓氏贵州鱼龙(Guizhouic thyosasaurus tangae),后经尚庆华等(2008)研究后认为,该种应该归属于萨斯特鱼龙。前人定名的亚洲杯椎鱼龙(Cy,bospondylus asiaticus)、优美盘江龙(Panjiang sasaurus)是该种的后出名。
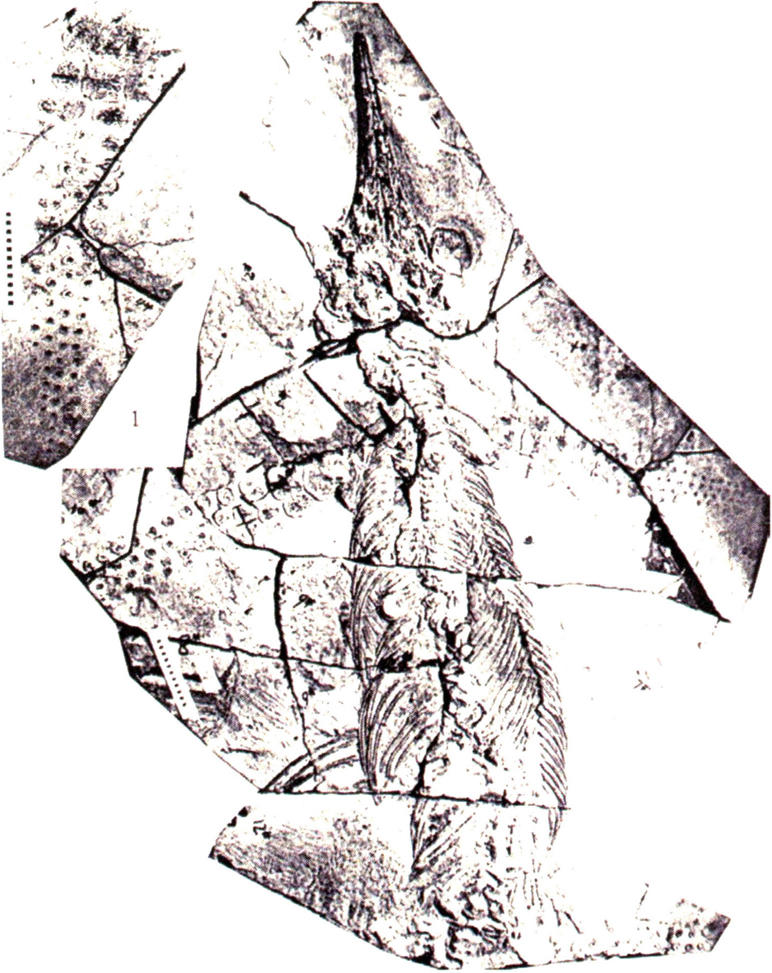
邓氏萨斯特鱼龙正型标本

邓氏萨斯特鱼龙(长600厘米)

萨斯特鱼龙头骨

萨斯特鱼龙与粗菊石(长450厘米)

萨斯特鱼龙(长560厘米)

萨斯特鱼龙(长700厘米)

萨斯特鱼龙生态复原图
关岭鱼龙
Guanlingsaurus
关岭鱼龙仅描述一个种,即梁氏关岭鱼龙(Guanlingsaurus liangae),产于贵州省关岭县新铺镇附近晚三叠世早期(卡尼期)法郎组瓦窑段。
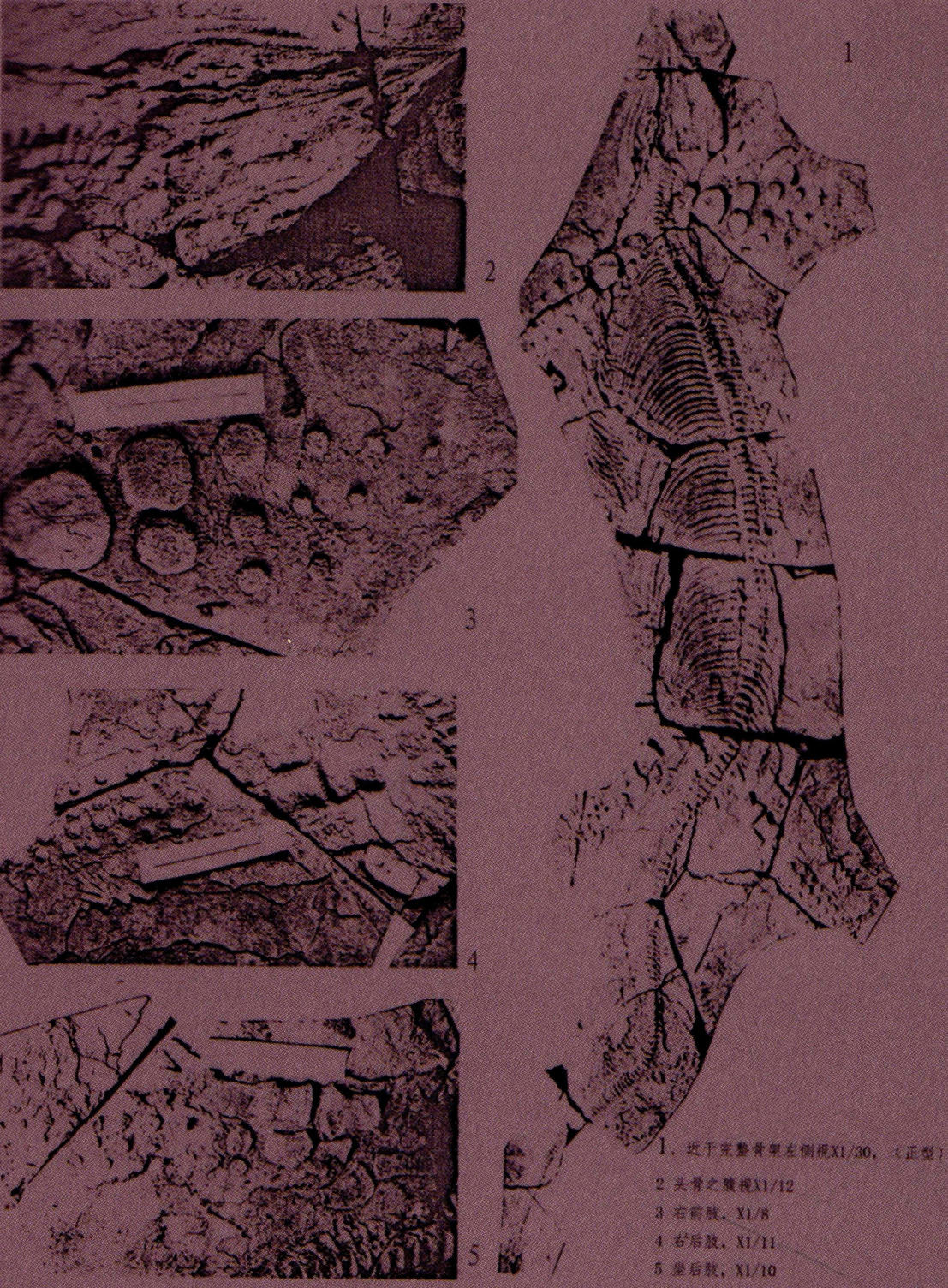
为一大型的鱼龙类,体长6米以上。头骨腹视为宽三角形,长略大于宽,下颌联合极短,左右翼骨联合将颅底封盖。颈椎较长,具短的劲肋,肋头宽而平,自近端向远端逐渐收缩变细,略弯曲,神经棘低,荐前椎约80节左右,椎体短,为双凹型,中部略下凹,横突低。肋骨较长,单头型,肋头扁而较宽平,肋骨中部有一自近端向远端延伸的浅沟。具有腹肋。尾部较长,荐椎和尾椎分节之和超过140节。尾椎椎体短,高大于长,为明显的双凹型,横突弱,“人字骨”(或脉脊、脉弧)“丫”形,神经棘低。尾椎后部明显向下弯曲。
肩甲骨小而扁,似三角形和扇形,桡骨近方形,宽略大于长,肱骨、尺骨、腕骨、掌骨和指骨均缩短特化为圆形,桡、尺骨间间隙极小,精密排列。腕骨间间隙和掌骨间间隙都比较大。指骨3列,疏松排列,指骨列间间隙和指骨节间间隙都很大。
肠骨较细,耻骨比较发育,坐骨扁平;股骨和胫骨为两端略扩展,中部稍许收缩的粗棒状;腓骨为内侧微凹的半椭圆形。胫、腓骨间间隙较小,紧密排列。跗骨、蹠骨和趾骨亦缩短特化为圆形。跗骨间间隙和蹠骨间间隙都较大。趾骨4列,疏松排列,趾骨列间间隙和趾骨节间间隙都很大。前肢大于后肢,前、后肢鳍脚均较长,属长鳍型。
李承(2009~2013)对该标本观察研究,并结合其他标本对比研究后指出:
一、关岭鱼龙与北美晚三叠世的shonisaurus近似,两者构成姐妹群;二、发现关岭鱼龙具尾弯,说明其已有很强的游泳能力,完全能够适应远洋遨游。同时,将尾弯出现的时间从侏罗纪早期提前到晚三叠世早期。

梁氏关岭鱼龙(长650厘米)

萨斯特鱼龙(长455厘米)
海龙类
Thalathosaurids

海龙类是三叠纪海生爬行动物中一个重要门类,虽已知其属种不多,但是形态迥异,生态分异开度很大,海龙体长3米以上,其尾部特别长,可以超过整个身体的一半。四肢则保留着陆地祖先动物的原始形态。
海龙虽然形体比较大,但其身体尤其是四肢的形态并未完全适应水中生活。它们的游泳能力有限,不具远洋生活能力,只能在近岸浅海中活动。
海龙主要依靠尾部的侧向波动推动身体前进,死后尾部在化石中常侧向保存。海龙的四肢,依然适合陆地行走,海龙的头骨分为长吻和短吻两种类型,吻都主要为前颌骨组成。鼻孔位置靠后,上颞孔缩小甚至消失,下颞弓不完整。枕部强烈向前凹,枕脊存在,枕骨位于下颌关节处之前,下颌骨有显著的冠状突。
海龙可能是全球分布最多的类群,在古特提斯海洋的西部和东部,以及古太平洋的东岸均有分布。
晚近的研究表明,海龙可能是双孔类的基干类群,也是三叠纪海洋中一个相当重要的成员。迄今为止,海龙已知种类才区区10来种,但形态各异,生态分异度极大。按其形态可分为谜龙科(Askeptosauridae)、海龙科(Thalattosaur.dae),两者的显著差异在于颈部。谜龙科的成员颈椎超过10枚,长度大于头长,而海龙科的成员颈椎一般少于7枚,颈部不太明显,目前在贵州已发现4属6种海龙。

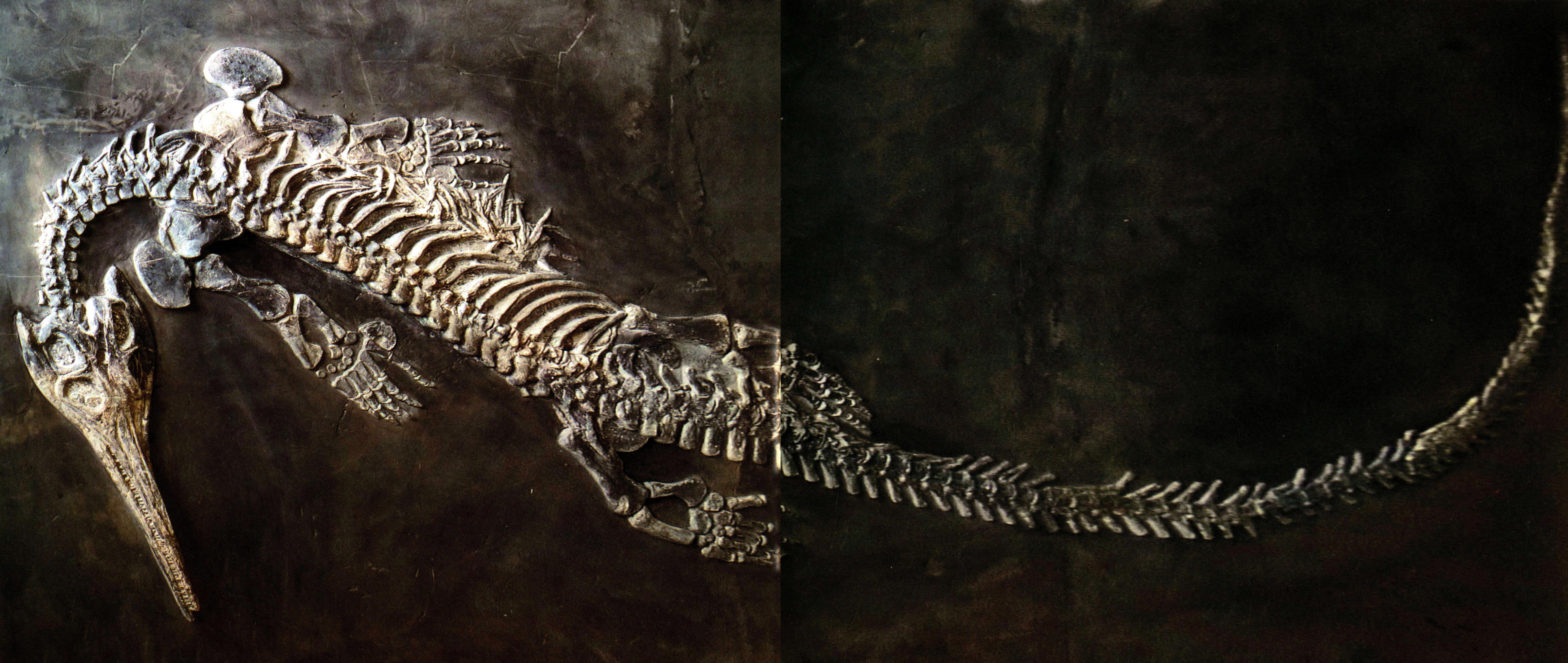
海龙

海龙与粗菊石(大龙体长140厘米,小龙体长80厘米)
海龙(长380厘米)
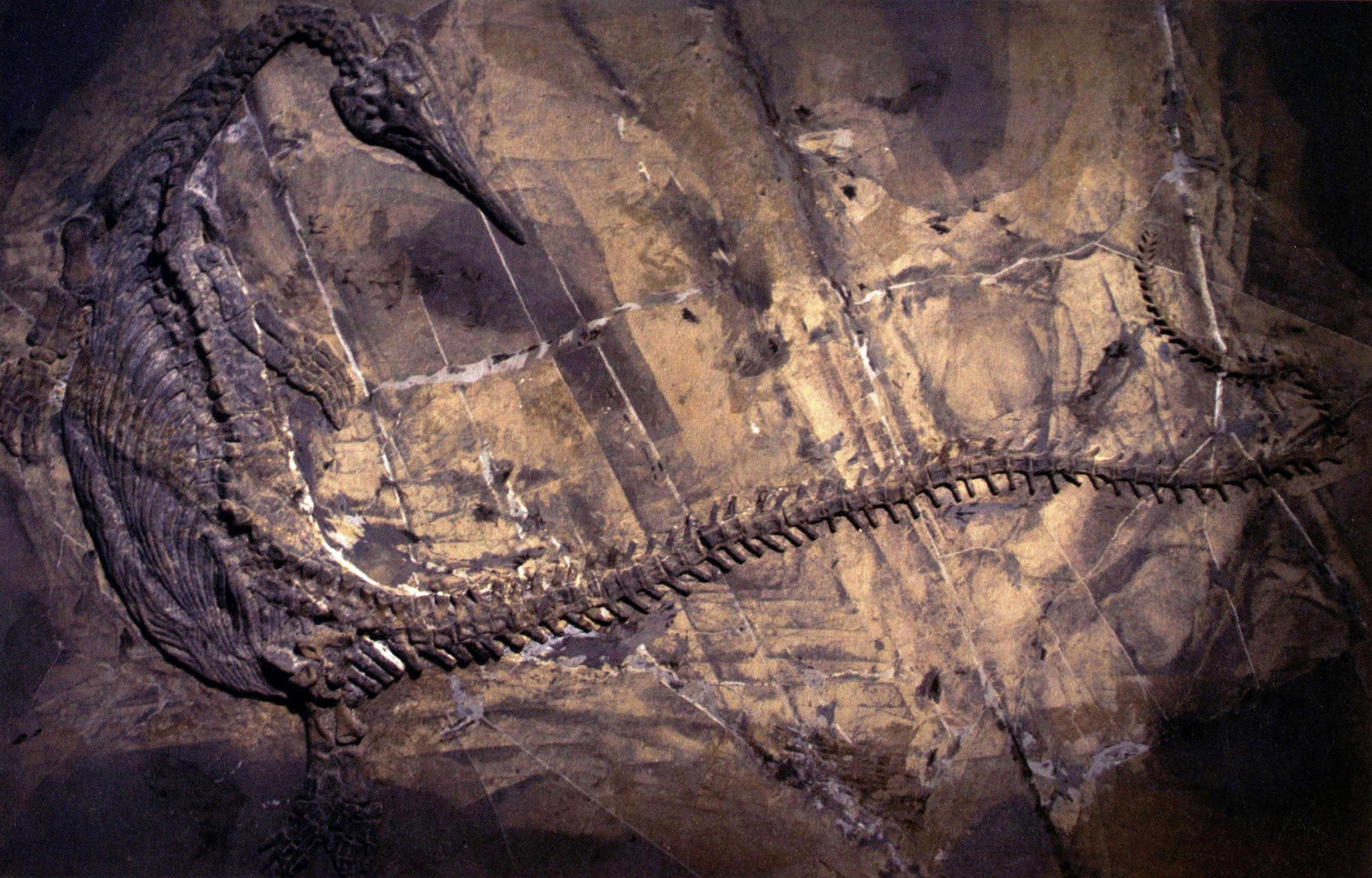
海龙(未定种,长460厘米)

海龙(大龙长170厘米,小龙长50厘米)

孙氏新铺龙(长128厘米)
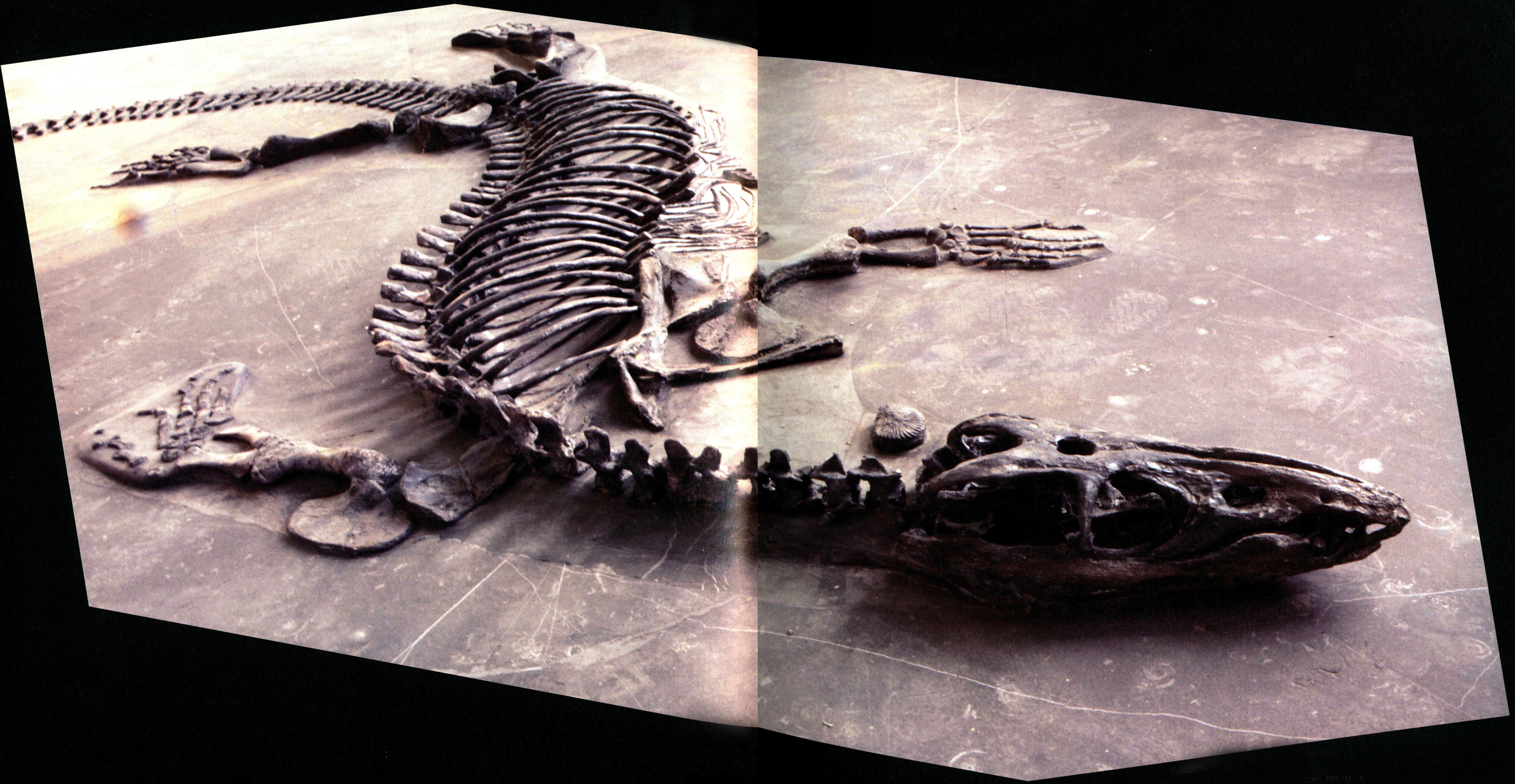
新中国龙(长520厘米)

海龙(未定属,长230厘米)

长216厘米

黄果树安顺龙(长110厘米)
一、谜龙亚类

乌沙安顺龙(长260厘米)
安顺龙
Anshunsaurus
安顺龙是我国第一个报道的海龙类。目前已发表描述了两种,即黄果树安顺龙(A.huangguoshuense)和乌沙安顺龙(A.wushaense),前者产于关岭县新铺镇一带晚三叠世早期法郎组瓦窑段,后者产于兴义市乌沙镇一带中三叠纪晚期法郎组竹竿坡段。
安顺龙已发现的标本从几十厘米到300厘米以上。其最显著特点是伸长的吻部,很长的脖子以及长长的尾巴,颈椎15~16枚,背椎超过23枚,荐椎2枚,尾椎数目在50枚以上。安顺龙头骨扁平,吻部长度大约为头长之半。牙齿为圆锥形同型齿,比较尖锐,颚面无齿。肩胛骨宽而低,X锁骨凹面关节,肩锁骨发起为T型。四肢发达,各个骨骼边缘都比较圆滑,缺乏显著的突起,基本保持脊膜动物的普通趾式。但黄果树安顺龙第五指,通常多一个指节,而乌沙安顺龙第四指失去一个指节。
安顺龙的上下颌牙齿比较尖锐,这种牙齿不适合压碎带壳的动物,推测其依靠捕食中小型鱼类为主。安顺龙的身体很长,四肢虽然发达但相对于身体较小,即便有一定的陆地活动能力,也不会多么敏捷,其上岸的主要目的可能是产卵或者晒太阳。四肢由于较小且不特化,不可能在游泳中起推进作用,走动主要靠尾部及身体的侧向波动。这种走动方式难以保持持续的高速,为了获取食物维持生存,它可能采取伏击方式捕食。

黄果树安顺龙(长200厘米)
贫齿龙
Miodentosaurus
目前仅见一个种,即短吻贫齿龙(Miodendentosaurus brevis),产于贵州省关岭县新铺镇附近法郎组瓦窑段,地质时代为晚三叠世早期(卡尼克中期)。
短吻贫齿龙的特征是:个体较大,全长超过400厘米,头长可达33厘米,吻直且极短,前额骨沿前骨中央有一隆嵴,上颌在前颌骨有6枚圆锥形齿,无上颌骨齿,头骨顶面松果孔大且很前位,头骨腭面的锄骨和翼骨的无齿,下颌齿骨齿都集中在前端且至多不超过6枚,上隅骨腹侧在后方1/3多突然加宽,向锁骨走端比近端窄得多,后肢趾式2、3、4、5、5。

短吻贫齿龙头骨(左为背视,右为腹视)

短吻贫齿龙(长390厘米)
二、海龙亚类
新铺龙
Xinpusaurus
新铺龙是已知最繁盛的海龙类,虽然化石只产于关岭新铺一带三叠世早期法郎组瓦窑段,但已有标本发现,新铺龙目前包括两个有效种,即孙氏新铺龙(Xinpusaurus sun yin effa)和巴毛林新铺龙(X.bamaolinensis),二者的区别在于吻部形态,孙氏种的吻部短,巴毛林种的前上颌骨强烈前伸,大大超过下颌的长端。
新铺龙一般体长1米多,最大个体长超过2米,吻部比较突出,两边呈锐角相交,齿列前面的牙齿较大,为圆锥形,后面的牙齿圆钝,呈纽扣状。头骨腭面上的梨骨与翼骨同样长有牙齿,梨骨齿稍大,为圆钝的球基状,翼骨上则排列着几列钝的小齿。新铺龙的颈部短,只有大约5枚颈椎;尾部长,尾椎大约90枚,神经棘比较发达,为高的宽板状,肩胛骨而高,骨与腓骨显著加宽,四肢较短。
新铺龙应该可以在水中及陆地活动,在水中游动可能主要靠尾部侧向摆动,四肢起到舵的作用,主要是控制方向。推测它们也生活在近岸的浅海中,因为这种环境食物最丰富。根据其牙齿形态,推测它们能够处理带壳的无脊椎动物(如双壳类),而食物的繁盛可以解释新铺龙的繁盛。

孙氏新铺龙
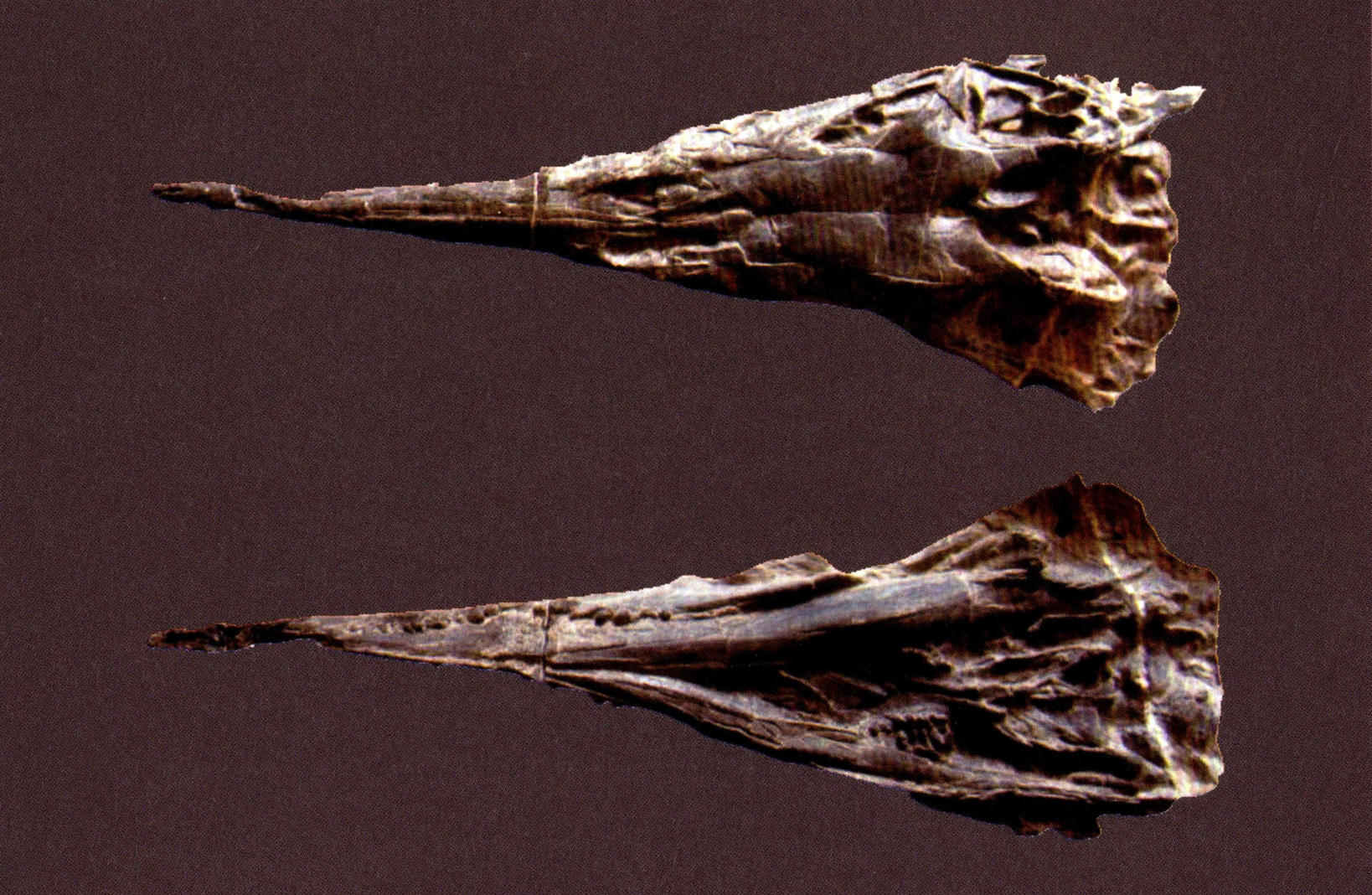
巴毛林新铺龙头骨(上为背视,下为腹视)
凹棘龙
Concavispina
凹棘龙只有一个种,即双列齿凹棘龙(Concavispina biseridens)。
标本产于贵州省关岭县新铺镇一带晚三叠世法郎组瓦窑段。顾名思义是上颌具有两列齿经棘背缘具凹口的海生爬行海龙类。
凹棘龙主要特征是体型较大,标本骨架长度为364厘米。其中不完整的头骨长50厘米,是目前已知最大的海龙类龙骨,上颌骨前端有两列交错排列的较钝的牙齿,后部无齿,背椎神经棘低,不超过前后长度的两倍,下宽上窄,背缘形成V形缺口,肩胛骨低宽,股骨骨干轻微收缩。
凹棘龙无疑可以归入海龙目,上颞孔偏小甚至消失,下颞弓缺失,头部眶后骨与后额骨愈合,方轭骨缺失,腓骨扩展,尾部长的扁平,其脊椎神经脊下宽上窄,背缘有V形缺口,这个独特征未见于其他海龙。而且其上颌骨具有两列牙齿在海龙中也是独有的,凹棘龙没有明显的颈部,额骨前侧突接近,明显属于海龙科。

双列齿凹棘龙(侧视)
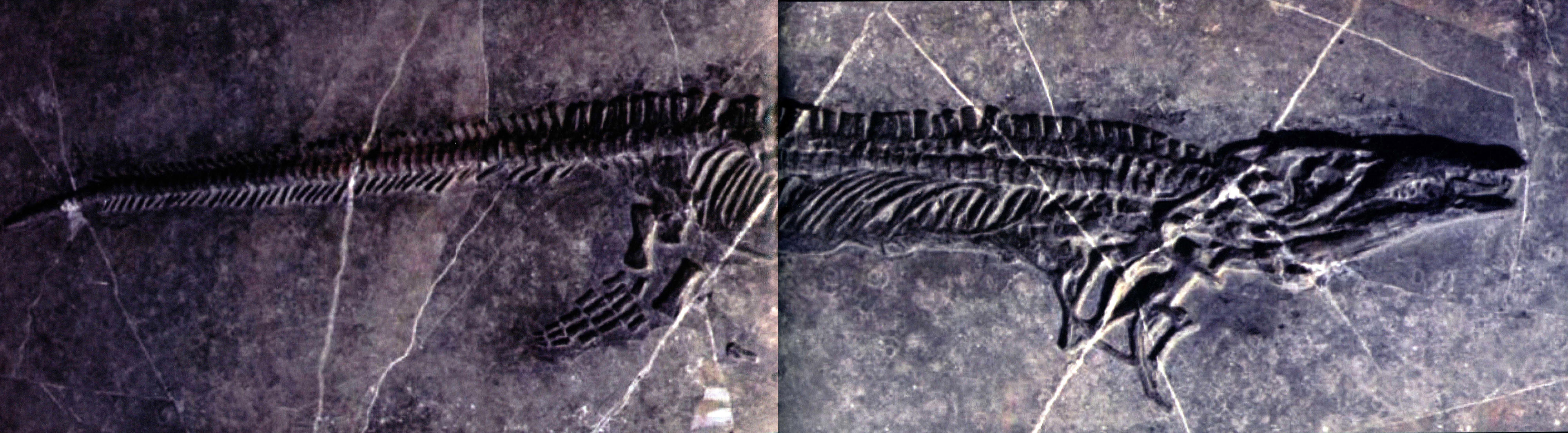
双列齿凹棘龙(长121厘米)
原龙类
Protorosaurians

东方恐头龙生态复原图
原龙类是早期初龙型爬行动物的一支,头骨两侧各有一个典型的上颞孔。原龙类开始于二叠纪,并延续到侏罗纪早期,这个类群的属种不多,以陆生动物为主,贵州发现的原龙类可分为海生及陆生两大类。
一、海生原龙类
贵州已发现的海生原龙类为长颈龙科的属种。长颈龙分布于中生代早期的特提斯洋近岸地区,包括欧洲、以色列和中国西南一带。长颈龙堪称有史以来最奇特的脊椎动物之一,这种史前爬行动物的整个颈部的长度超过体长的一半,颈椎纤细并极度拉长,具有发达的颈肋,一个颈肋相当于2~3节颈椎的长度。贵州已描述的长颈龙样化石包括两个属。
东方恐头龙(长330厘米)

海生原龙类(未定名,长151厘米)

蛇颈龙
东方恐头龙
Dinocephalosaurus orientalis
东方恐头龙产于盘县新民乡羊圈地区中三叠统早期(安泥期)关岭组第二段中上部。
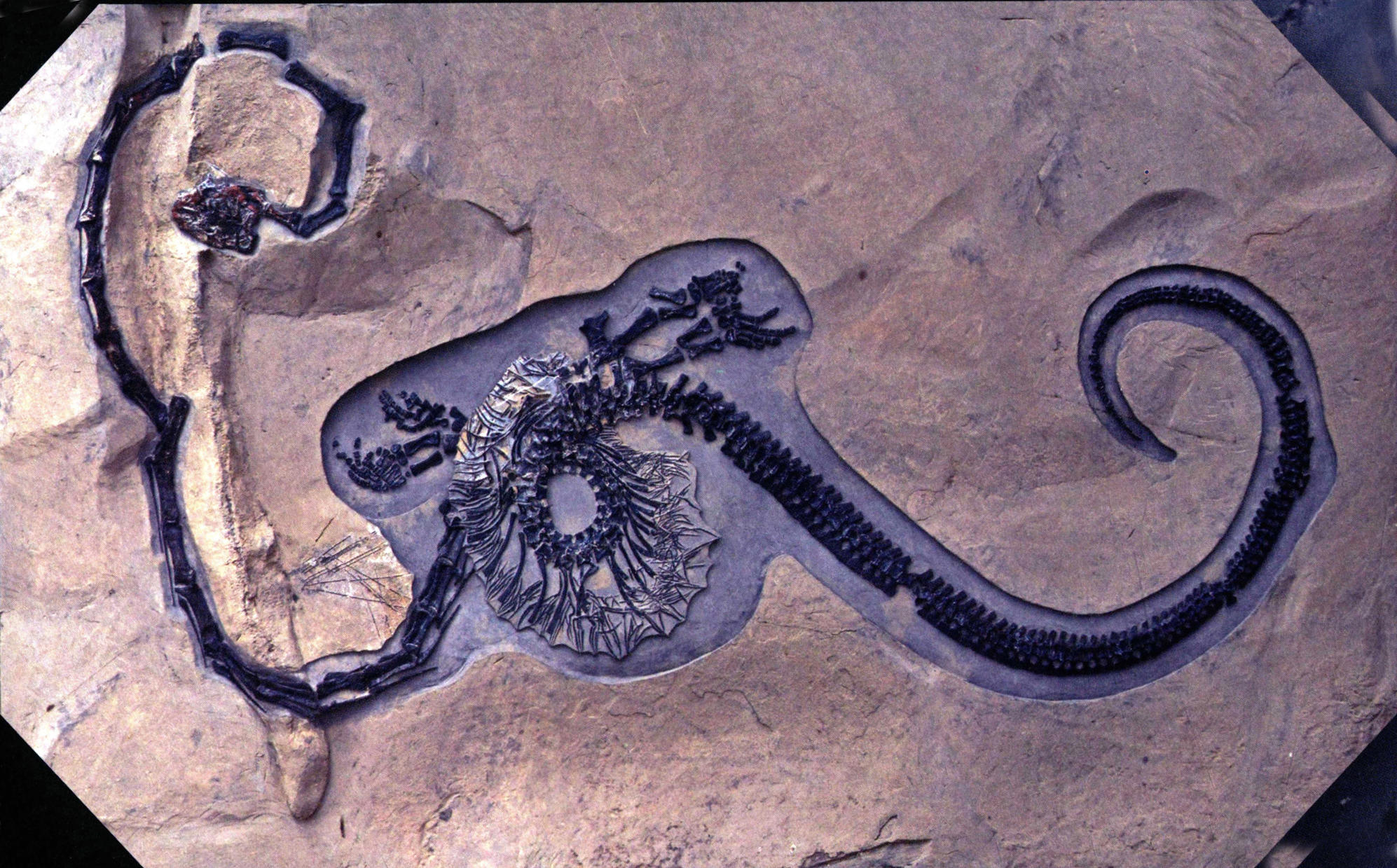
东方恐头龙的原意是“东特提斯地区具有恐怖头骨的爬行动物”。东方恐头龙具有一条极度伸长的脖子,颈部长1.7米,大大超过仅有1米的躯干长度,显然它和长颈龙结构相似——椎体伸长,神经弓不发育,都有相似的颈肋。但二者颈部的伸长方式存在差别,长颈龙的颈部仅有12节颈椎,每节颈椎都极度拉长;恐头龙单枚颈椎的长度不及前者,但颈椎数目大致在25枚以上。

东方恐头龙(长520厘米)
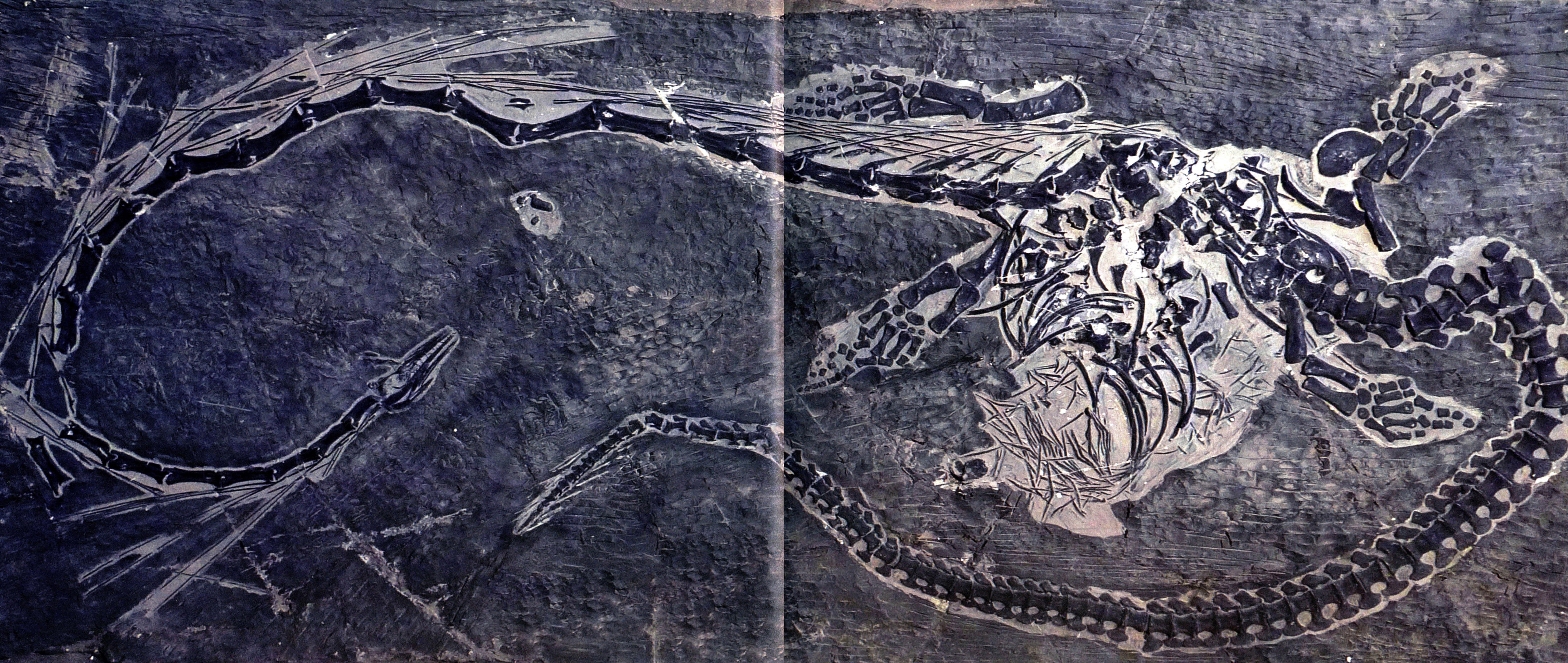
东方恐头龙(长530厘米)
长颈龙(未定种)
Tanystropheus sp.
现有记述的第二种海生原龙类标本采自贵州省兴义市中三叠统法郎组竹竿坡,是一个保存不完整的化石骨架,包括完整的躯干部,前肢和5枚连续保存的后部颈椎。骨架保存长度只有51厘米,骨骼细小,说明它是一个幼年个体。颈椎的神经棘不发育,椎体极度拉长。长宽比例明显地大于东方恐头龙的比例,而与欧洲产的长颈龙类相似。例如最后一节颈椎的长与宽之比,兴义标本是4.43,东方恐头龙是2.1,欧洲的长颈龙是4.39。因此,兴义标本被看作是长颈龙属的一个未定种。
由于身体结构细长而脆弱,长颈龙的化石大都保存不好,几乎没有完整的骨架。标本的局限增加了人们认识该类群的难度。时至今日,长颈龙类的形态功能学研究乃是古生物学领域的热点和难点之一,众说纷纭,莫衷一是。虽然历史上长颈龙类曾被看作是陆生动物,或根据幼年标本上的“三尖齿”认为其幼年个体是陆生动物,捕食昆虫,成年后进入海洋生活。但它们的腕骨为简单的圆形,骨化程度弱,表明长颈龙应是终身海生爬行动物。
中生代海生爬行动物通过两种方式演化出超长的颈部。第一种方式见于鳍龙类,在三叠纪的幻龙、纯信龙和侏罗纪、白垩纪的蛇颈龙中,动物的每节颈椎都很短,颈肋也很短,而颈椎的数目极大,最多可以达到70枚。第二种方式见于海生的原龙类,颈椎和颈肋都很长,但颈椎数目相对很少。两种结构迥异的长颈,如果它们的运动方式不一样,那么后者更有可能是僵直的。由于长颈龙的颈肋特别细长,达到2~3节椎体的长度,这样每两节椎体之间的链接处都被2~3对,或者说4~6根颈肋束缚着。如果这些颈肋不具有一定程度的柔韧性,就意味着这样一条长颈无法弯曲,便无法灵活运动。研究者在讨论东方恐头龙的颈部功能时曾提出过一种“吞吸”的捕食方式,即对于一条无法灵活运动的长脖子而言,利用颈部肌肉的突然收缩控制细长的颈肋,通过食道体积的猛烈扩张而产生“吸力”,从而捕捉到猎物。

东方恐头龙
二、陆生原龙类
至2014年底仅描述一属一种,即富源巨颈龙(Macrocnemus fuyuanensis)——中国南方海相三叠系中第一件真正意义上的陆生爬行动物化石。
陆生原龙类体长1米左右,头骨三角形,口内布满细密的牙齿;四肢纤细,前肢短,后肢长,快速运动时可以两足行走。我国发现了一件巨颈龙化石,其体内残留着贵州龙的遗骸,化石显示它的大腿曾经骨折,但伤口已经愈合。它是否在海滨浅水里捕食贵州龙,却不料受到其他海怪的偷袭,一番殊死搏斗后大难不死?
在云南东部和贵州西部发现的巨颈龙,是在瑞士圣乔治山地区以外的首次发现。这一发现,不仅为颇具争议的竹竿坡时代归附于中三叠世拉丁期提供了新的化石证据,同时也揭示出该地区存在一个未知的三叠纪陆生爬行动物群。这个陆生爬行动物群,可能源于中三叠世早期,至拉丁期达到成熟阶段。同时还表明,中生代早期特提斯洋北缘的岛屿和岛链,是多种爬行动物特别是陆生爬行动物迁移的重要通道。
标本产于贵州省兴义市乌沙镇与云南省富源县大箐村交界处中三叠世晚期法郎组竹竿坡段。
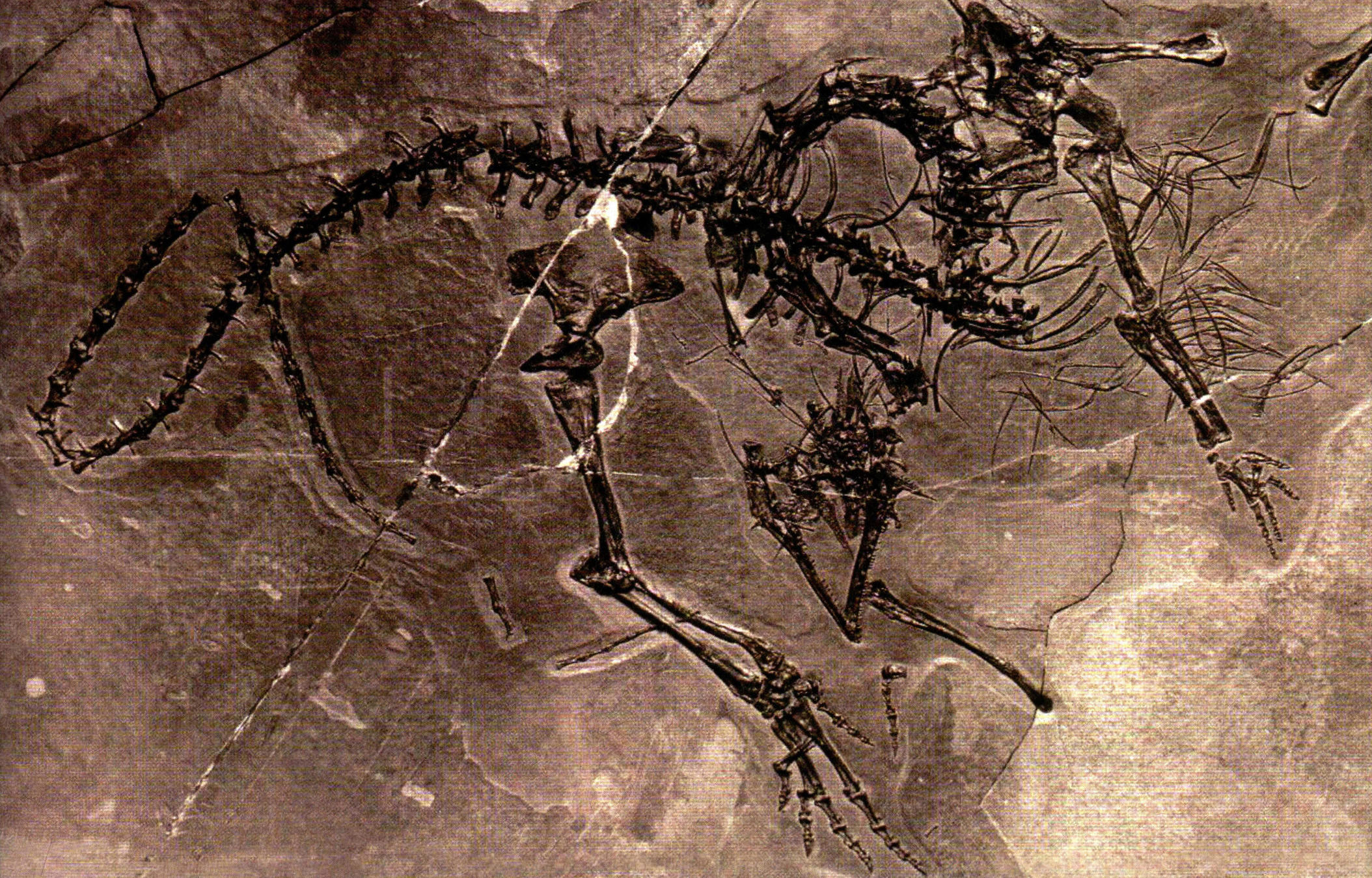
体内残留着贵州龙遗骸的巨颈龙

富源巨颈龙生态复原图
主龙类
Archosaurians

主龙类是爬行动物中一个庞大而复杂的类群,中生代的恐龙(包括鸟类)、翼龙以及现代的鳄鱼都是其重要成员。这三个类群之外的主龙形类爬行动物,通常被称作“槽齿类”,也就是本文所称的主龙类,其共同特征是外形细长,鼻骨长、下颌细长,槽生齿等。主龙类化石材料虽不丰富,但赋存于地球上各个大陆,对生物学和地球历史的研究具有极重要的意义。长期以来,“槽齿类”化石在世界各地均被发现于陆相沉积地层中,晚近,中国古生物工作者首次发现了海相地层中保存的“槽齿类”完整骨架,并在化石上找到许多动物水生的特征,打破了古生物学界长期以来“槽齿类没有进入海洋”的论断。
云贵地区中三叠统发现的主龙类化石计有2属2种。
混形黔鳄
Qianosuchus mixtus li
混形黔鳄产于贵州省盘县中三叠世早期关岭组第二段的上部。
混形黔鳄得名是因其兼具海生和陆生特征并在贵州发现,一条卷曲状保存的,体长超过3米的大型爬行动物化石。
混形黔鳄中等体型,前颌骨鼻骨支细长,9枚前颌骨齿;外鼻孔后位,长度大于头骨上的其他开孔,后缘位于眶前孔前缘之前,上、下缘分别由鼻骨、前颌骨和上颌骨构成;下颌孔半卵圆形;第2~7颈椎的神经棘前后加宽,每节神经棘顶部覆盖有5枚小骨板;尾椎神经棘高,远高于相应的人字骨,且至少三倍于其椎体高度;颈肋纤细,超过相应椎体长度的三倍;肩胛骨宽大,呈斧头状。
混形黔鳄以低矮的前颌骨、9枚匕首状的前颌骨齿、后位且狭长的外鼻孔(由鼻骨、前颌骨及上颌骨构成,长于头骨上的任何其他开孔)、半卵圆形的下颌孔、细长的颈肋、斧头状的肩胛骨以及高大的尾椎神经棘等特征区别于其他初龙类。
确定混形黔鳄兼具海生和陆生特征的依据有下述几点:(1)化石保存十分完整,只有部分尾椎缺失,所有骨骼全部相互关联。这是动物死亡后未经搬运而原地保存埋藏的有力证据。原地埋藏的完整骨架,产于海相碳酸盐岩地层中,同层中产有海生鱼类和无脊椎动物化石(如双壳、腕足类等)。(2)该动物骨骼具有一些明显海生特征,如宽大的肩带骨骼,即盘状的、薄的肩胛骨和乌喙骨。这一特征在诸多海生爬行动物类群如鳍龙类、鱼龙类和苍龙类中都极为常见。另外巨大而狭长并接近头骨顶部位的鼻孔、纤细的颈肋等特征也都是海生爬行动物中最常见到的。最能反映混形黔鳄适应海生生活的特征是其尾部结构。混形黔鳄的尾椎具有高度发达的神经棘和人字骨,表明其尾部极为侧扁。在爬行动物中,最主要的游泳方式是依靠躯体的侧向摆动推动自身前进,而在绝大多数海生爬行动物,例如鱼龙类、海龙类、沧龙以及某些鳄类中,更主要的是依靠尾部的侧向摆动。在这些类群中,尾部都是侧扁的。(3)研究人员惊异地发现这一标本的腰带和后肢骨骼明显地适应陆地生活,它后肢强壮,腰带类似于蜥臀类恐龙,完全没有任何水生动物特征。很显然,混形黔鳄在陆地和近岸浅海中都具有很强的运动能力,它的后肢比前肢更强壮,在陆地上可以像恐龙一样用四足或两足行走;在海水中,它主要通过尾部的强烈摆动推动身体前进。不论在陆地还是海洋,混形黔鳄都是可怕的杀手,它的嘴里布满匕首状、边缘带有锯齿的牙齿,可以轻而易举地刺穿和撕碎任何猎物,包括同时期的鱼类、幻龙类、海龙类、原龙类,也许还有鱼龙类等。

混形黔鳄(这是世界上第一具海生初龙类化石)

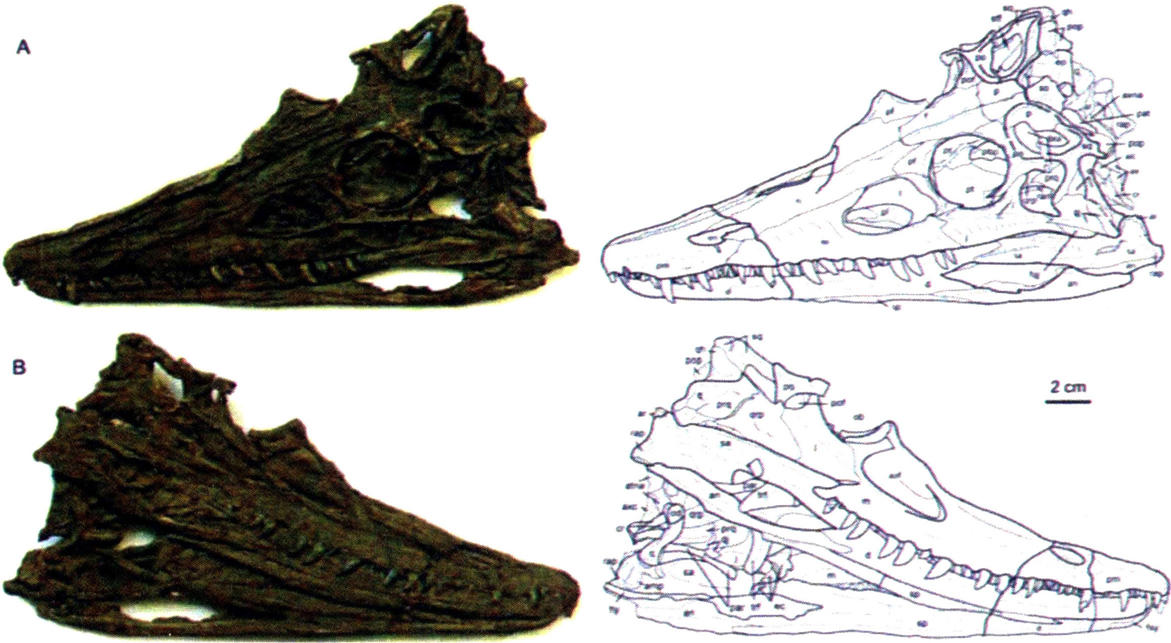
混形黔鳄头骨(AB:侧面,CD:腭面)

混形黔鳄复原图
混形黔鳄骨骼复原
主龙类(长298厘米)
富源滇东鳄
Diandongosuchus fuyuanensis
富源滇东鳄产于云南省富源县十八连山乡大箐村附近,中三叠世晚期(拉丁期)法郎组竹竿坡段,是中等的主龙类,与其他主龙类的主要区别是:眶前部长度约为眶后长度的2.5倍;前颌骨具鼻孔窝,且鼻骨支向后延伸超越外鼻孔;前颌骨齿9枚上颌骨不进入外鼻孔;轭骨表面具明显纵脊,前肢明显宽于后肢,末端插入方轭骨之下;上颞孔位于颞窝后部;颈椎骨板前端窄而深凹;乌喙孔大,紧邻肩胛骨;坐骨中部扩展;蹠骨中第四蹠骨最长。

富源滇东鳄生态复原图
与混形黔鳄相比,富源滇东鳄显示出更加倾向于陆地生活的骨骼特征。尽管如此,滇东鳄有一些特征,例如位置相对靠后的外鼻孔、表面具纹饰的颅顶骨骼以及数目多的前额骨牙齿等,在纯粹的陆生初龙类中并不常见,而见于包括混形黔鳄在内的半水生或有水生倾向的主龙类。其头骨特征以及标本腹内的鱼骨化石碎片表明滇东鳄与现代咸水鳄具有类似的生活模式,营海岸附近的两栖生活。
富源滇东鳄头骨(A:顶面,B:腭面)

富源滇东鳄正形标本
龙龟类
龙龟(Saurosphargis)是欧洲西特提斯动物群中一类罕见而奇异的海生动物。
1903年,一位法国古生物学者在研究一块极为残破的、仅存的几根扁平肋骨和零星骨板的化石标本时,命名了这个几乎只由几根肋骨代表的神秘史前动物为“Saurosphargis”(中文翻译为龙龟)。此后的100多年间再没有Saurosphargis新化石的发现。更糟的是,它的正型标本也在第二次世界大战中丢失,仅有一具模型留存。直至2003年,西方学者才再次发现了一个极为破碎的标本,这些七零八落的散碎骨块虽然具备某些类似“Saurosphargis”的特征,但是仍旧无法解析这种动物整体形象。可以说“Saurosphargis”自发现命名至今一个多世纪中,从来就是“有名无实”且不为人们重视的“怪物”。
2012年,中国科学院古脊椎动物与古人类研究与浙江自然博物馆首次在云贵地区三叠纪海相地层中发现此类动物较为完整的化石,并命名为中国龙龟(Sinosaurosphargis yunguiensis)。中国龙龟是极具特征化的海生双孔类,最显著的特征是头骨上颞孔几近于封闭而下颞孔完全开放,这种情况在水生爬行动物中极为罕见,仅见于极个别的楯齿龙;体表覆盖着无数真皮骨板组成的背甲。与楯齿龙不同的是,这些小骨板不仅分布于躯干部,而且同样覆盖着四肢和尾椎、弓状的尖齿以及宽大有短粗凸起的肋状骨。
作为鳍龙类的姐妹群,龙龟类本身分为两个支系,包括具有甲壳的中国龙龟和不具甲壳的大头龙。
系统发育分析显示出中国龙龟与海龙类和鳍龙类具有更加密切的亲缘关系。

黔大头龙(产于贵州省盘县新民乡羊圈村附近中三叠世早期关岭组二段)

黔大头龙(产于贵州省盘县新民乡羊圈村附近中三叠世早期关岭组二段)
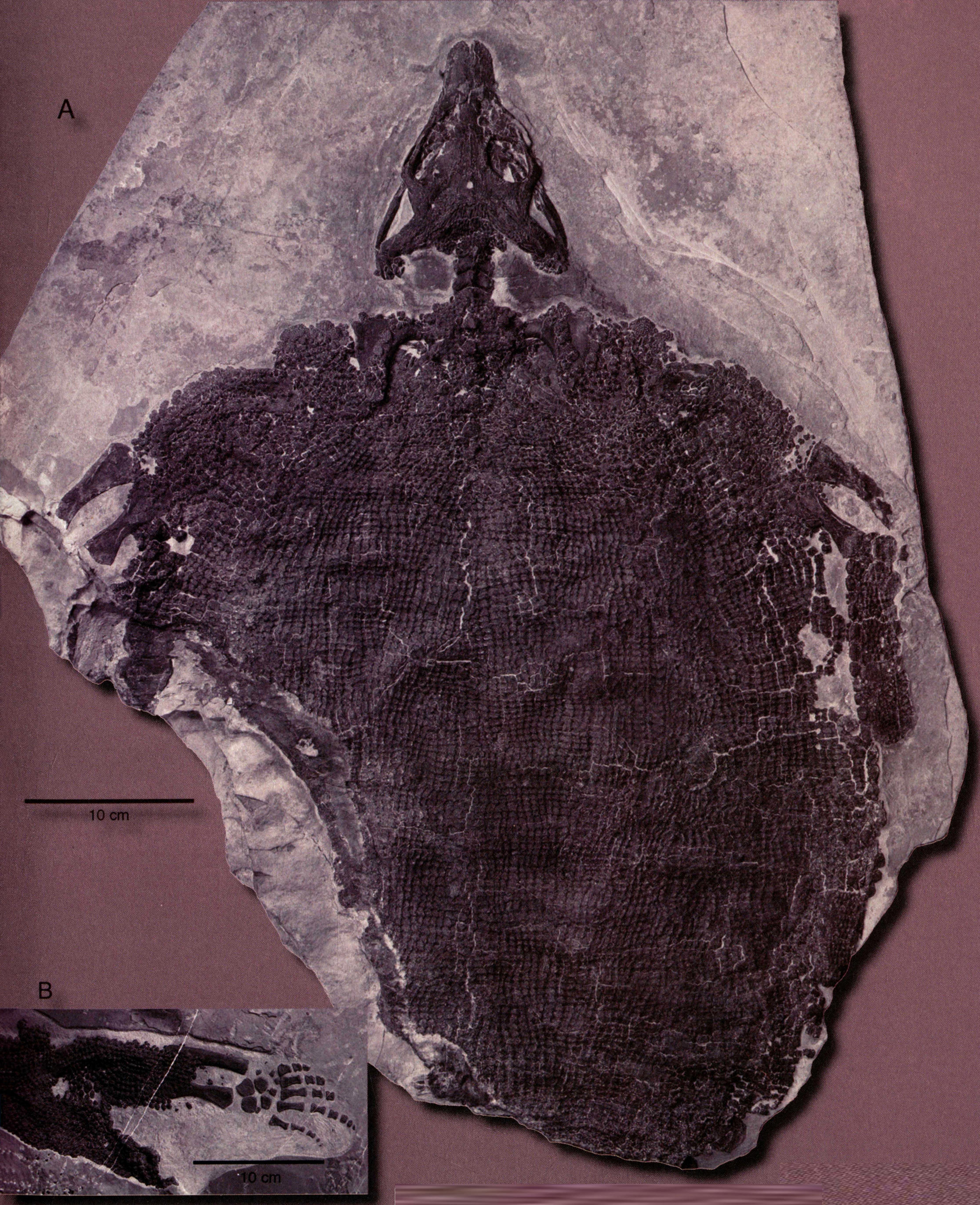
云贵中国龙龟(长172厘米)。产于贵州省盘县、云南省罗平县等地中三叠世早期关岭组二段。
龟鳖类
Testudines

龟鳖类生态复原图
龟鳖类是爬行动物中最为特化的一类,身体宽短,背腹具甲。这种沉重的装甲是一种被动性的保护,大多数种类的颈和四肢、尾部可以在一定程度上缩进甲内。甲可分为背甲和腹甲,有的种类尚有下缘甲,连接背腹甲。长期以来,将龟鳖类归于无孔类,但最新的形态学和分子生物学研究认为,龟鳖类是双孔类爬行动物。
半甲齿龟
Odontochelys semitestacea li et al
半甲齿龟是世界上最早最原始的龟类化石,发现于贵州省关岭县新铺镇沙星村晚三叠世早期法郎组瓦窑段。
半甲齿龟具有细密的牙齿以及雏形状态的甲壳结构。龟类腹甲的形成远远早于背甲,当腹部甲壳已进化到与今天的龟类相差无几时,背部的甲壳才开始出现。其背甲的形成与肋骨和脊椎的特化有显著关系,此外,半甲齿龟的身体结构及其化石产生地点的地质情况表明,这种原始龟类很可能生活于海窪或者河流三角地带,适应于水生环境的程度与今天的鳖相似。半甲齿龟不仅代表了龟类起源过程中的过渡阶段,而且首次从古生物的角度揭示了这一物种特殊身体结构的进化。
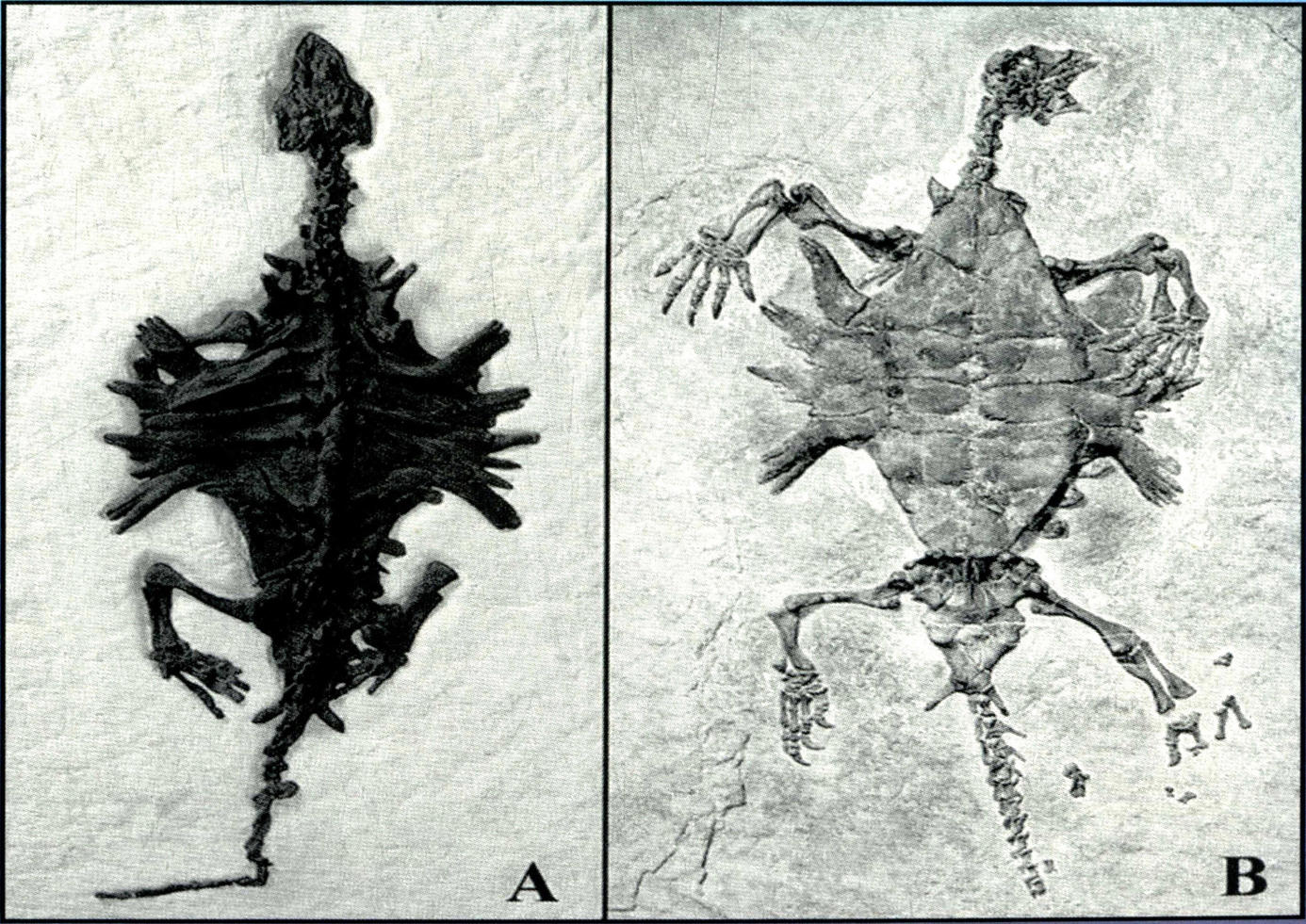
半甲齿龟